In Figure 1 we show the gating strategy for the isolation of hematopoietic stem cells from the mouse bone marrow and the layout of the plate for their ex vivo culture. Figure 1A shows the identification of the lymphocyte fraction in the SSC-A/FSC-A plot. Doublets were removed in the singlet gate followed by identification of live cells by the absence of DAPI signal. The LKS population, defined by lineage- Sca1+cKit+, was identified. This population is known to contain stem and progenitor cells. HSCs form around 5–10% of the cells in the LKS population and were identified by gating for CD150+CD48– population. Figure 1B represents the layout of the 96-well plate for ex vivo culture. Sorted HSCs were plated in different culture conditions:In this case, control and NR supplemented culture conditions. Whole bone marrow cells were also plated as single-color controls as described in the protocol. It is important to fill all surrounding wells with water to avoid evaporation of media from cell-containing wells. Moreover, as mentioned previously, marginal wells were avoided for cell culture because they are more susceptible to evaporation.
Figure 2 shows the measurement of mitochondrial membrane potential (ΔΨm) and mass in HSCs post culture. Figure 2A shows representative plots of TMRM levels (above) and green fluorescent mitochondrial stain (below) in HSCs cultured in control and NR supplemented conditions. NR treatment showed a clear increase in the TMRMlow population. Figure 2B shows the quantification from three independent samples. NR treatment significantly increased the proportion of cells in the TMRMlow gate and showed a significant lowering of TMRM fluorescence intensity. Mitochondrial mass (represented by green-fluorescence intensity) did not change upon NR supplementation. Additionally, we combined stem cell marker staining with mitochondrial staining post culture. Figure 2C shows the gating strategy to identify HSCs from lineage negative and LKS populations post culture in the two culture conditions. The TMRM and green fluorescent mitochondrial stain profile of these gated HSCs is seen in Figure 2D. Exposure to NR showed a significant increase in the %TMRMlow population and a significant decrease in the TMRM fluorescence intensity in gated HSCs. The green fluorescent mitochondrial stain green signal remained unchanged in the two conditions.
Figure 3 shows the measurement of mitochondrial membrane potential (ΔΨm) and mass in different human T cells: peripheral blood mononuclear cells (PBMCs) CD4+ and CD8+ T cells, as well as CD4+ and CD8+ tumor infiltrating lymphocytes (TILs) after the rapid expansion protocol (REP). Figure 3A shows representative plots of the TMRM levels (above) and green-fluorescent mitochondrial stain levels (below) of circulating (PBMC) and tumor-infiltrating (TIL) CD4+ and CD8+ T cells. TILs showed a clear increase in TMRM and green fluorescent mitochondrial stain signal compared to circulating T cells. Figure 3B shows the MFI quantification of TMRM and green fluorescent mitochondrial staining. TILs displayed higher TMRM and green fluorescent mitochondrial staining signals compared to PBMC-derived T cells. These data indicate that TILs have a distinguished metabolic profile with increased ΔΨm and mitochondrial mass.
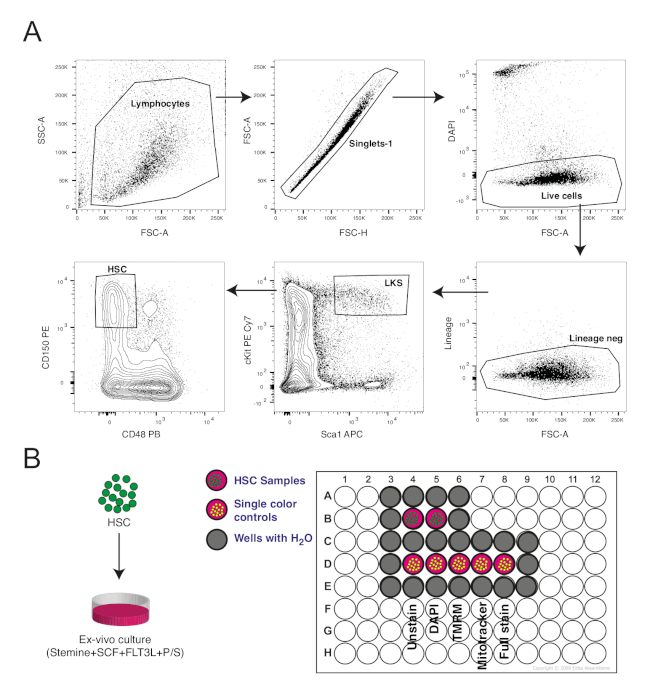
Figure 1: Isolation and culture of hematopoietic stem cells. (A) Gating strategy for isolation of hematopoietic stem cells (HSCs) based on cell surface markers. HSCs were identified as lineage- Sca1+ cKit+ (LKS) CD150+CD48–. (B) Design of 96 well plate put in culture. Please click here to view a larger version of this figure.
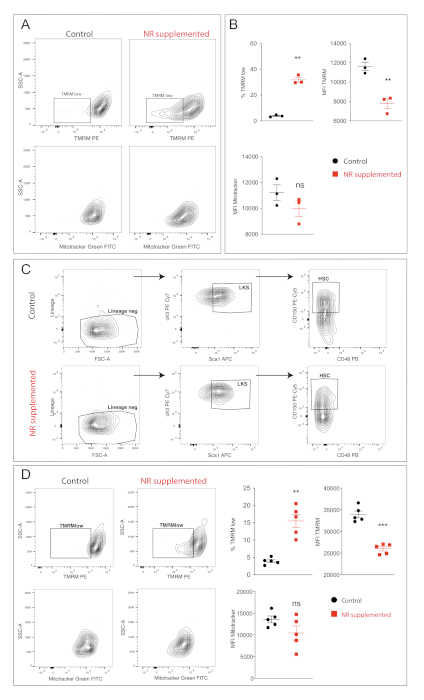
Figure 2: Mitochondrial profiles of HSCs. (A) FACS contour plot showing HSCs post culture in basal or NR supplemented conditions. TMRM (above) and green fluorescent mitochondrial stain (Mitotracker) (below) profiles are shown. (B) Quantification of TMRM and green fluorescent mitochondrial stain signal. NR supplementation resulted in a decrease in TMRM profile while maintaining the green fluorescent mitochondrial stain signal. (C) Contour plots showing identification of HSC population in control and NR supplemented conditions post culture. (D) Contour plots and quantification of TMRM and green fluorescent mitochondrial stain signal in phenotypic HSCs post culture. NR supplementation reduced the TMRM signal while the green fluorescent mitochondrial stain signal remained unchanged in HSCs. Student t test ***p < 0.001, ** p < 0.01, * p < 0.05, not significant > 0.05, error bars = SEM. Please click here to view a larger version of this figure.
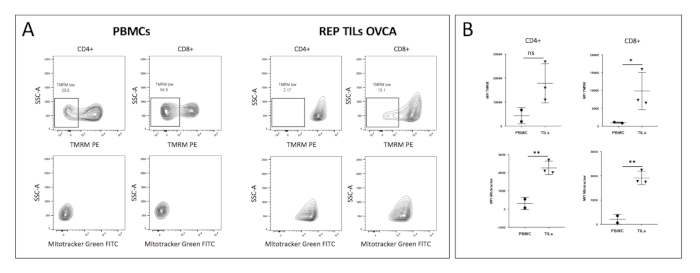
Figure 3: Mitochondrial profiles of human PBMCs and TILs: (A) FACS contour plot showing CD4+ and CD8+ freshly isolated from PBMCs or tumor-derived CD4+ and CD8+ post REP (rapid expansion protocol). TMRM (above) and green fluorescent mitochondrial stain (Mitotracker) (below) profiles are shown. (B) Quantification of TMRM and green fluorescent mitochondrial stain signal. TILs displayed lower mitochondrial activity and mass. Student t-test ***p < 0.001, **p < 0.01, *p < 0.05, not significant > 0.05, error bars = SD. Please click here to view a larger version of this figure.
| S.No | Antibody name | Working dilution |
| 1 | Streptavidin Tx red | 1/200 |
| 2 | Sca1 APC | 1/200 |
| 3 | Ckit PeCy7 | 1/100 |
| 4 | CD150 PE | 1/100 |
| 5 | CD48 PB | 1/100 |
| To be used only if stem cell and mitochondrial markers combined. | ||
| 6 | Streptavidin Pac orange | 1/200 |
| 7 | CD150 PE-Cy5 | 1/100 |
Table 1: Antibody dilutions.
