The figures describe protocols used to overcome the challenge of attaching high lipid and chitin-containing chorion Drosophila embryos (Table 1) to the glass slide surface for examination and experimentation. Utilizing the chrome alum gelatin slide-coating method shown in Figure 1, we enhanced the attachment of Drosophila embryos onto the surface of slides while the embryo pre-embedding method shown in Figure 2 allows for efficient inspection of the dynamic distribution of protein and RNA DmFKBP12/Calstabin in all four early development stages (i.e., syncytial blastoderm, cellular blastoderm, early and late gastrula of the Drosophila embryos). Figure 3 shows the protein distribution of antibody detected FKBP12 / Calstabin at the syncytial and cellular blastoderm stages. From these histochemical results, it can be seen that FKBP12 expression is less polarized in syncytial blastoderm (Panels A to H), and then begins to localize within the basement membrane underneath the trophectoderm epithelium to form a gradient with more expression found in the anterior than the posterior poles (Panels I to S). Eventually, the DmFKBP12/Calstabin protein becomes restricted to certain architectural components of the primitive embryonic tissues with less extracellular distribution than that observed in early and late gastrula embryonic stages (Figure 4). Interestingly, the above-described dynamic protein distribution is not accompanied by corresponding mRNA levels of DmFKBP12 (Figure 5), indicating protein expression to be the result of a still unknown post-transcription molecular mechanism. The described method of CAG slide-coating and pre-embedding will allow us to further probe the importance of DmFKBP12 in calcium signaling and unearth this still unknown molecular mechanism while combining other techniques such as micro-injection RNA interference19.
In this study, we develop a chrome alum gelatin slide-coating and pre-embedding method for histology, histochemical and immune-histological analysis of Drosophila embryos at different development stages. Using this pre-embedding method, the embryos are effectively collected for both cryo- and paraffin sectioning that allows for the analysis of longitudinal cross sections. This allowed us to describe the evolving expression patterns for this protein during decisive developmental stages, linking expression of the protein to development of key physiologic systems such as the gut, the brain, and the connecting nervous system. By employing these methods, we were able to investigate the dynamic distributions of RyR regulating protein and mRNA FKBP12/Calstabin during the development of Drosophila embryos at definitive stages, including the syncytial and cellular blastoderm, and early and late gastrulation stages. Our data showed that the FKBP12/Calstabin protein and its RNA can be detected very early in the syncytial blastoderm. As the embryo develops, FKBP12 becomes more densely distributed in the basal cell layer underneath the epithelium of the blastoderm. The FKBP12/Calstabin protein then dynamically strengthens its expression and differentiates its distribution into embryonic muscle-containing tissues, including the stomodaeum, stomodaeum and posterior midgut rudiment. These distribution changes from the beginning of blastoderm to late gastrulation stages are also described in the developing neuronal system including neuroblasts, ventral nerve, supraesophageal ganglion and brain. While these dynamic changes in FKBP12/Calstabin hint at its importance in development, they are not reflected in mRNA levels of the protein, hinting at a yet unknown regulator of protein expression that remains to be identified.
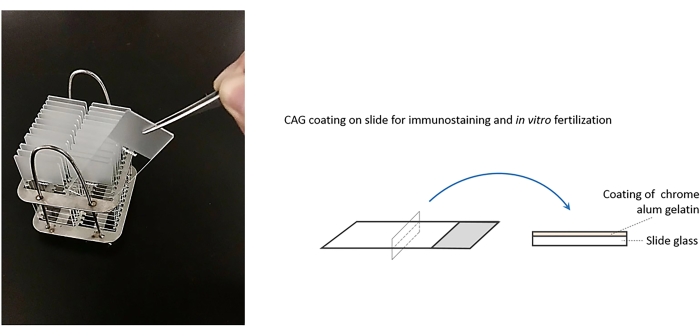
Figure 1: Sketch of chrome alum gelatin (CAG)coating method for paraffin-sections used in immunology and RNA in-situ hybridization to visualize the distribution of DmFKBP12 protein and mRNA in Drosophila embryos. Please click here to view a larger version of this figure.
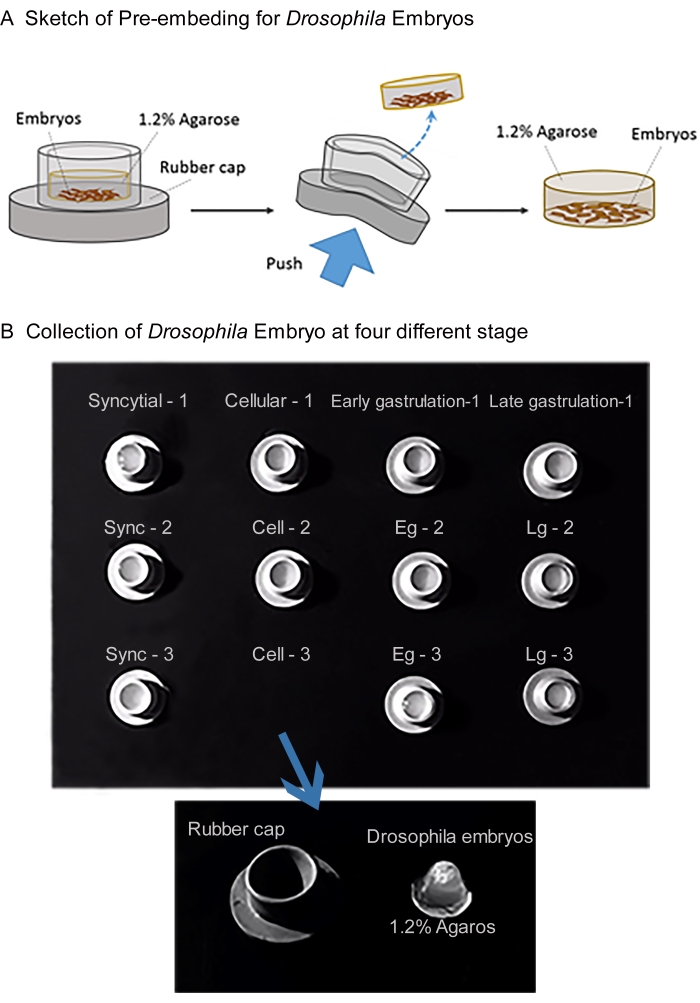
Figure 2: Sketch of pre-embedding method for expression profile of FKBP12 and DmFKBP12 in developing Drosophila embryos. Please click here to view a larger version of this figure.
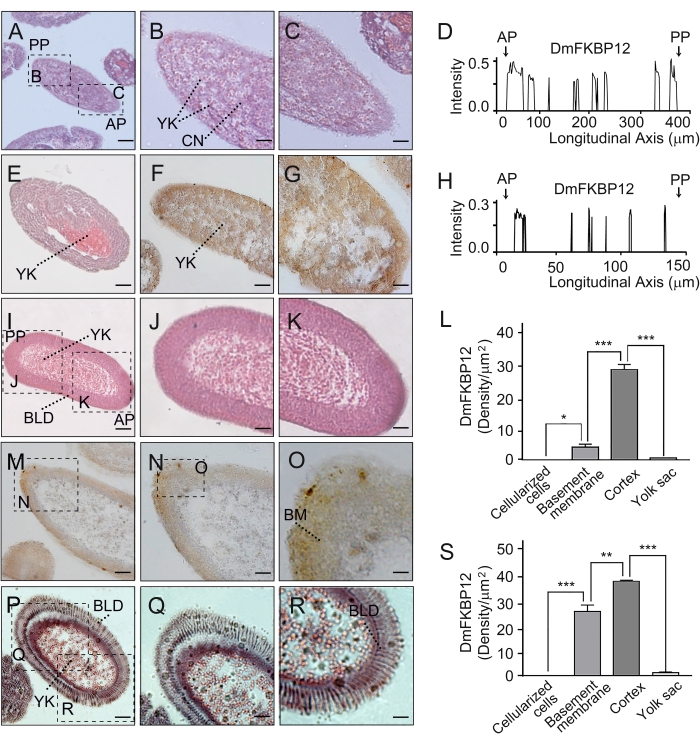
Figure 3: Expression profile of DmFKBP12 protein in syncytial and cellular blastoderms of the Drosophila embryo. (A-D) H&E staining of the syncytial blastoderm. Quantification of DmFKBP12 distribution in early syncytial blastoderm is presented in D. (E-G) Panels present early syncytial blastoderm in which nuclei are under division. Quantification of DmFKBP12 distribution in late syncytial blastoderm are presented in H. (I-L) H&E staining of the cellular blastoderm. The qualification of DmFKBP12 distribution in late cellular blastoderm is presented in L. (M-S) Panels exhibit the distribution of DmFKBP12 protein in the cellular blastoderm. O shows the larger views of M-N. (P-R) Panels present earlier cellular blastoderm shown in M-O. Quantifications of DmFKBP12 distribution in early cellular blastoderm is presented in S. Scale bar for A, E, I, M and P is 60 µm, for B, C, F, J, K, N, Q and R is 40 µm, and for G, O is 20 µm. AP, anterior pole of egg; YK, yolk; CN, cleavage nucleus; PP, posterior pole of egg; BLD, blastoderm, nuclei and cell; BM, Basement membrane. Data are expressed as mean± SEM. * p < 0.05, ** p < 0.01, *** p < 0.001. The data quantifying was carried out with ImageJ. Firstly, the brown products were remained in the image along with removing all other colors. Secondly, the brown image was changed black-and-white and calculate its density for quantification consequently. Please click here to view a larger version of this figure.
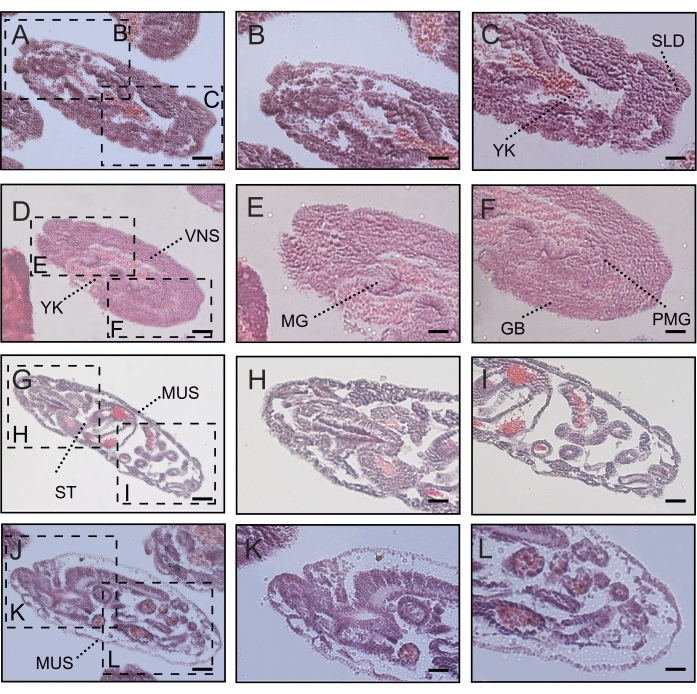
Figure 4: Expression profile of DmFKBP12 protein in early and late gastrulation stages of the Drosophila embryo. A, D and G show H&E staining of Drosophila embryo at the gastrulation stage. B, C, E and F display the distribution of DmFKBP12 at the early gastrulation stage. H, I, and J-L display the distribution of DmFKBP12 at the late gastrulation stage. Quantifications of DmFKBP12 distribution in early gastrulae are presented in M. Scale bar for A, D, E and G is 80 µm, for B, C, F, H, I and J-L is 40 µm. PP, posterior pole of egg; NBL, neuroblasts; ST, stomodaeum; YK, yolk; PMG, posterior midgut rudiment; BR, brain, supraesophageal ganglion; VNS, ventral nervous system; PV, proventriculus; MUS, muscle; MGC, midgut caecum; AMG, anterior midgut rudipighian; MMG, middle midgut; TR, trachea. Data are expressed as mean± SEM. ns, no significant; ** p < 0.01, *** p < 0.001. The data quantifying was carried out with ImageJ 1.50d by remaining product-brown and changing to black-and-white image, and following by density calculation of black-and-white as described in the previous figure legend. Please click here to view a larger version of this figure.
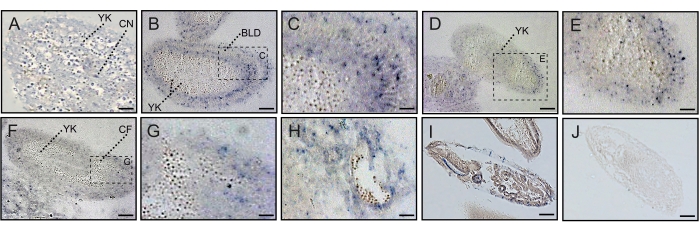
Figure 5: In-situ localization of DmFKBP12 mRNA during embryonic development of the Drosophila embryo. A shows in-situ localization of DmFKBP12 mRNA at the syncytial blastoderm stage when FKBP12 mRNA was expressed in the cytoplasm of the embryo. B and C show in-situ localization of DmFKBP12 mRNA at the cellular blastoderm stage when FKBP12 mRNA was expressed in the inner limits of blastoderm cells. D and E exhibit in-situ localization of FKBP12 mRNA at the early gastrulation stage. During this stage, it is mainly localized in the developing gut. F, G and H show in-situ localization of FKBP12 mRNA at the late gastrulation stage, when the mRNA is expressed in the muscle and gut. I is positive control of DmFKBP12 mRNA. J is negative control of DmFKBP12 mRNA. Scale bar for A, C, E, G and I is 5 µm, for B, D, F, I and J is 10 µm. YK, yolk; CN, cleavage nucleus; BLD, blastoderm, nuclei and cells; CF, anterior oblique cleft, cephalic furrow. Please click here to view a larger version of this figure.
| Embryonic stages | 0-2 hours (%) | 2-3 hours (%) | 3-12 hours (%) | 12-24 hours (%) |
| Syncytial blastoderm | 90.8 | 0 | 0 | 0 |
| Cellular blastoderm | 6.9 | 91.54 | 0 | 0 |
| Early gastrula | 2.3 | 4.23 | 91.1 | 0 |
| Late gastrula | 0 | 4.23 | 8.93 | 93.3 |
Table 1: Embryo collection of different stages in development of Drosophila melanogaster
