Denuded prophase I-arrested oocytes of 21-day-old C57/BL6 mice were injected with a reporter mix containing mRNA encoding the Ypet reporter fused to the 3' UTR of IL-7 and mRNA encoding mCherry. YFP and mCherry expression was recorded in 39 oocytes, of which 30 were matured, and 9 were arrested in prophase I as a negative control. Three maturing oocytes were excluded for analysis because they either had a delayed GVBD (N=2) or moved in the dish during the recording (N=1). Figure 3 shows mCherry and YFP expression in prophase I and maturing oocytes. Figure 4 shows YFP expression of maturing oocytes corrected for plateaued mCherry expression (averaged mCherry expression in the last 10 time-points) to correct for the injected volume. Translation rates of the reporter were measured by curve-fitting (linear regression) the YFP/mCherry values in prophase I and in maturing oocytes during the first 0-2 h or 8-10 h after cilostamide release (Figure 5A). The accumulation of the reporter does not follow a linear pattern, as indicated by a significant difference in translation rates between 0-2 h and 8-10 h after cilostamide release (p<0.0001; Figure 5B). Therefore, these results indicate activation of IL-7 translation during oocyte meiotic maturation.
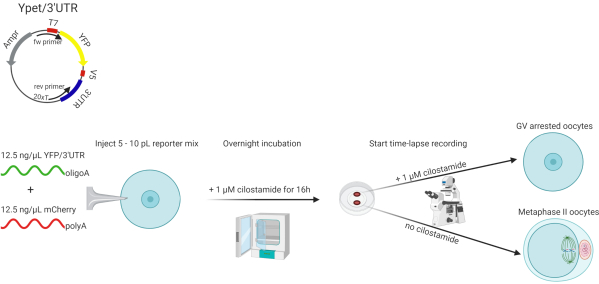
Figure 1: Schematic overview of the experimental procedure. Oligoadenylated Ypet/3' UTR and polyadenylated mRNA encoding mCherry are micro-injected into denuded oocytes of 21-day-old C57/BL6 mice. Oocytes are pre-incubated for 16 h in cilostamide containing maturation medium to allow the mCherry signal to reach a plateau. After pre-incubation, a time-lapse recording is started where oocytes are either kept in medium with cilostamide to create a prophase I-arrested control group or released in cilostamide-free medium to mature. Abbreviations: UTR = untranslated region; YFP = yellow fluorescent protein; fw primer = forward primer; rev primer = reverse primer; Ampr = ampicillin resistance; polyA = polyadenyl; oligoA = oligoadenyl; GV = germinal vesicle. Please click here to view a larger version of this figure.
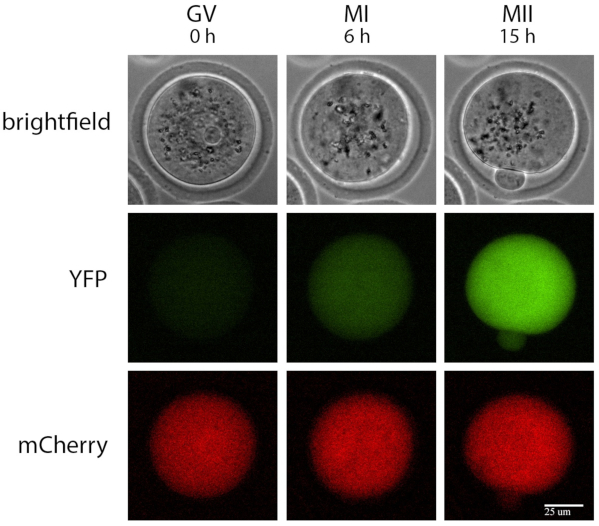
Figure 2: Example of a single oocyte time-lapse recording. Brightfield, YFP, and mCherry recordings of a single oocyte injected with mRNAs encoding Ypet/Interleukin-7 3' UTR and polyadenylated mCherry at prophase I, MI (6 h after cilostamide release), and MII (15 h after cilostamide release). Scale bar = 25 µm. Abbreviations: YFP = yellow fluorescent protein; GV = germinal vesicle; MI = metaphase I; MII = metaphase II. Please click here to view a larger version of this figure.
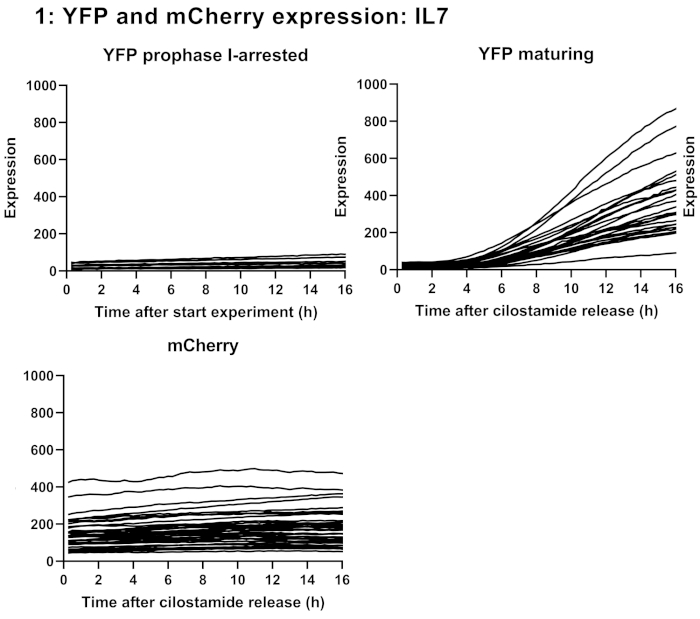
Figure 3: YFP and mCherry signals recorded by time-lapse microscopy. YFP and mCherry signals of oocytes injected with oligoadenylated Ypet/IL7 3' UTR and polyadenylated mRNA encoding mCherry. Oocytes were either kept in medium with cilostamide to generate a prophase I-arrested control group (N=9) or released in cilostamide-free medium to allow for maturation (N=30). Data are individual oocyte measurements. Abbreviations: IL-7 = interleukin-7; YFP = yellow fluorescent protein. Please click here to view a larger version of this figure.

Figure 4: YFP signal corrected for co-injected mCherry level. YFP signals of prophase I-arrested and maturing oocytes were corrected for injected volume by dividing the YFP signal by the average mCherry signal of the last 10 time-points. Individual YFP/mCherry ratios for (A) prophase I-arrested oocytes and (B) maturing oocytes and mean ± standard error of the mean YFP/mCherry ratios of (C) prophase I-arrested and (D) maturating oocytes. Abbreviations: YFP = yellow fluorescent protein; GVBD = germinal vesicle breakdown. Please click here to view a larger version of this figure.
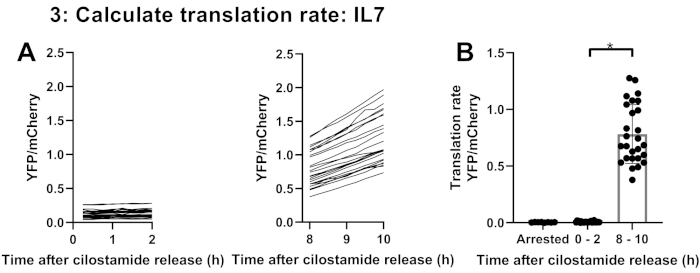
Figure 5: Calculated translation rates at 0-2 h and 8-10 h of maturation. (A) Yellow fluorescent protein (YFP) signals of single oocytes corrected for mCherry (YFP/mCherry) at 0-2 h or 8-10 h after cilostamide release and (B) translation rates (mean ± standard error of the mean) as calculated by curve-fitting (linear regression) the YFP/mCherry values at 0-2 h and 8-10 h after cilostamide release. Data were analyzed using the unpaired two-tailed t-test. *p < 0.0001. Please click here to view a larger version of this figure.

Figure 6: Example of repression of translation: Oosp2. Re-analyzed data of an experiment where oocytes were injected with Ypet-Oosp2 3′ UTR and polyadenylated mRNA encoding mCherry. YFP signals of prophase I-arrested (N=63) and maturing oocytes (N=72) were corrected for injected volume by dividing the YFP signal by the average mCherry signal of the last 10 time-points. YFP and mCherry expression data were obtained using a Xenon Arc lamp, unlike the IL-7 experiment where an LED light source was used. Data represent the mean ± standard error of the mean of individual oocyte measurements and were previously published in Luong et al.5. Abbreviations: YFP = yellow fluorescent protein; Oosp2 = oocyte secreted protein 2; UTR = untranslated region; IL-7 = interleukin-7; LED= light-emitting diode; GVBD = germinal vesicle breakdown. Please click here to view a larger version of this figure.
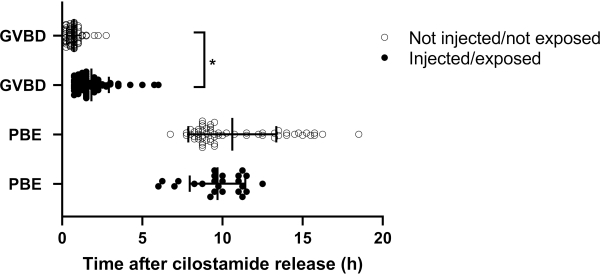
Figure 7: Effect of micro-injection and fluorescence exposure on timing of GVBD and PBE. Timing of GVBD and PBE of oocytes that were either micro-injected and exposed to fluorescence (injected), or not-injected and not exposed to fluorescence (not-injected). Data are individual oocyte measurements. Data were analyzed using the unpaired two-tailed t-test. *p<0.001. Abbreviations: GVBD = germinal vesicle breakdown; PBE= polar body extrusion. Please click here to view a larger version of this figure.
| Basic oocyte collection medium | ||
| Component | For 500 mL | |
| HEPES modified Minimum Essential Medium Eagle | 7.1 g | |
| Sodium bicarbonate | 252 mg | |
| Sodium pyruvate | 1.15 mL | |
| Penicillin/Streptomycin 100x | 5 mL | |
| Ultrapure distilled water (Invitrogen, 10977-015) | Up to 500 mL | |
| Maturation medium | ||
| Component | For 500 mL | |
| MEM alpha 1x | Up to 500 mL | |
| Sodium pyruvate | 1.15 mL | |
| Penicillin/Streptomycin 100x | 5 mL | |
Table 1: Preparation of media. List of components that need to be added to prepare basic oocyte collection medium and oocyte maturation medium.
| Sequence | |||||
| Ypet/Interleukin-7 3’ UTR | GAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTG GCTAGTTAAGCTTGGTACCGAGCTCGGATCCACCGGTCGCCACCATGGTGAGCAAAGGCGA AGAGCTGTTCACCGGCGTGGTGCCCATCCTGGTGGAGCTGGACGGCGACGTGAACGGCC ACAAGTTCAGCGTGAGCGGCGAGGGCGAGGGCGACGCCACCTACGGCAAGCTGACCCTG AAGCTGCTGTGCACCACCGGCAAGCTGCCCGTGCCCTGGCCCACCCTGGTGACCACCCTG GGCTACGGCGTGCAGTGCTTCGCCCGGTACCCCGACCACATGAAGCAGCACGACTTCTTCA AGAGCGCCATGCCCGAGGGCTACGTGCAGGAGCGGACCATCTTCTTCAAGGACGACGGCAA CTACAAGACCCGGGCCGAGGTGAAGTTCGAGGGCGACACCCTGGTGAACCGGATCGAGCTGA AGGGCATCGACTTCAAGGAGGACGGCAACATCCTGGGCCACAAGCTGGAGTACAACTACAAC AGCCACAACGTGTACATCACCGCCGACAAGCAGAAGAACGGCATCAAGGCCAACTTCAAGAT CCGGCACAACATCGAGGACGGCGGCGTGCAGCTGGCCGACCACTACCAGCAGAACACCCC CATCGGCGACGGCCCCGTGCTGCTGCCCGACAACCACTACCTGAGCTACCAGAGCGCCCTG TTCAAGGACCCCAACGAGAAGCGGGACCACATGGTGCTGCTGGAGTTCCTGACCGCCGCC GGCATCACCGAGGGCATGAACGAGCTCTATAAGAGATCTTTCGAAGGTAAGCCTATCCCTAA CCCTCTCCTCGGTCTCGATTCTACGCGTACCGGTCATCATCACCATCACCATTGAACAGGAC ATGTAGTAACAACCTCCAAGAATCTACTGGTTCATATACTTGGAGAGGTTGAAACCCTTCCAG AAGTTCCTGGATGCCTCCTGCTCAAATAAGCCAAGCAGCTGAGAAATCTACAGTGAGGTATG AGATGATGGACACAGAAATGCAGCTGACTGCTGCCGTCAGCATATACATATAAAGATATATCAA CTATACAGATTTTTGTAATGCAATCATGTCAACTGCAATGCTTTTAAAACCGTTCCAAATGTTTC TAACACTACAAAGTCTACAAAAAGCAAGGCTATGAAGATTCAGAGTCACCACTGTTTTCTTAGC AAAATGATGGTATGGTTAAACATTCATTGGTGAACCACTGGGGGAGTGGAACTGTCCTGTTTTAG ACTGGAGATACTGGAGGGCTCACGGTGATGGATAATGCTCTTGAAAACAAGAGTCTATCTTAAAGC AGCAGCAAAAAGAAGCTTAAGGCACTTAAGGCATCAACAAATGTAGTTAAATATGAATGTATAACA CATAACTTCAGTAAAGAGCATAGCAGATATTTTTAAATAAAAGTATTTTTAAAGATAGAAATGCACTTAT TCCAAAGATACTGAACCTTAGTATTCAGTCGCTTTTGACACTTGTGTATAATAAAGCTTATATAACTGAA TTTTCAATTTGAAAAGTATATTTTTAAAAGAATAATATATGCTAGACTTTTAATTAATGTATATGTTTAATTT TGGCATTCTGTCTGTCTCTCTGTCTCTCTCTCTCTCTCTCTCTCTCTCTACCTATCTATCTATATATATA ATTTTCATATACTACCAATTGCGTACTTTGGATAGTGTCTCTTTTTAACCTAAATGACCTTTATTAACAC TGTCAGGTTCCCTTACTCTCGAGAGTGTTCATTGCTGCACTGTCATTTGATCCCAGTTTTATTGAACAC ATATCCTTTAACACACTCACGTCCAGATTTAGCAGGAGACTAGGACCCTATAACTTTGTTAAGAGAGAA AACACTAATTTCTTGTTTTATAGTAGGGTCTTATTCGTATCTAAGGCAGGCTAGGATTGCAGACATGAGC CAATATGCTTAATTAGAAACATTCTTTTTATGTTAAACTCATGTCTTTTACAAGATGCCTACATATATCCTAT GTATATGCCTGTTTAAATCCTTTTTTGTAAGGTCTGCTGTCTTCCTTCAGTTGTAATGGAAAGAAACACTA TGTTGTAGAGGCCAAATTTCTGAAAGTGATAAGGGTTTGCTTGTACTGAATTCTCATTCTCCTTGCTTT TTCCAGCCACGTGAGCATCTAGCTATCTATACGCTGGATGTATTTGACCGATGCCTGCTCCACTGGCAC ATTGCATGTGTGGTAGCCATGCCTTCTTGCTTCTCCTTTTCCCCAACCCCTATAATGCTCTACTCAGTGG TACAGATAGCTGGGATTATCACAATTTTGAGAGAAACACCAATTGTTTAAAGTTTGTTTCATAATCACCATTT GCCCAGAAAACAGTTCTCTCAACTTGTTTGCAACATGTAATAATTTAAGAAACTCAATTTTGTTAATGGACTT TCGATAACTTCCTTAGATATCCCACATCTCCTACGTGTCAGTCCTTTGTCCTGAGGAACTGGTAAAATGGGTA AGCCCTTAGCTAGCGAACTGAAGGCATTCGCATGTGTAAGATAATCTCTATACCTGCAAGGCTGTCTGGAT GGCTCCCTACCAATATTGAACAATATTCTGATTTTGGCAAAATAAAGGATAATATTTT |
||||
| Forward primer | GAGAACCCACTGCTTAC | ||||
| Reverse primer | TTTTTTTTTTTTTTTTTTTTAAAATATTATCCTTTATTTTG CCAAAATC |
||||
Table 2: Example of reporter and primer sequences Sequence of YFP/IL7 3'UTR reporter and sequences of forward and reverse primers that were used to produce a linear PCR template for in vitro transcription.
Supplemental File: Yellow fluorescent protein (YFP) and mCherry time-lapse recording. YFP and mCherry time-lapse recordings of a single oocyte injected with mRNAs encoding Ypet/Interleukin-7 3' UTR and polyadenylated mCherry. YFP channel (Ex: S500/20 × 49057; Em: D535/30 m 47281), mCherry channel (Ex: 580/25 × 49829; Em: 632/60 m). The oocytes were recorded every 15 min for 16 h (7 frames/second). Please click here to download this file.
