All animal protocols and experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of the University of California Los Angeles.
1. Preparation of 58Fe-Tf
NOTE: The protocol uses 58Fe; however, an identical protocol can be used for 57Fe. Either isotope can be used and disposed of as a standard iron chemical without additional precautions.
- Dissolve 58Fe in 12 N HCl at 50 µL of HCl/mg of 58Fe.
- Add HCl to the metal in the glass vial supplied by the vendor, and replace the cap loosely. To dissolve the iron, warm the 58Fe/HCl solution to 60 °C for 1 h. If still not dissolved, leave the solution overnight at room temperature in the fume hood to dissolve.
NOTE: Dissolved 58Fe/HCl solution is yellowish-orange in color.
Fe3O4(s) + 8HCl(aq) → Fe(II)Cl2(aq) + 2Fe(III)Cl3(aq) + 4H2O
- Add HCl to the metal in the glass vial supplied by the vendor, and replace the cap loosely. To dissolve the iron, warm the 58Fe/HCl solution to 60 °C for 1 h. If still not dissolved, leave the solution overnight at room temperature in the fume hood to dissolve.
- Oxidize any remaining Fe(II)Cl2 to generate the Fe(III)Cl3 solution.
- Warm up the 58Fe/HCl solution to 60 °C with the cap off to facilitate oxidation.
- Add 1 µL of 35% H2O2 per 50 µL of 58Fe/HCl solution to further facilitate oxidation.
Fe(II)Cl2(aq) + O2 + 4HCl → 4Fe(III)Cl3(aq) + 2H2O
- Prepare the ferric chloride (58Fe(III)Cl3) solution.
- Leave the ferric chloride solution in the hood at 60 °C with the cap off to evaporate the sample.
NOTE: Evaporation may take between one and several days. - Reconstitute 58Fe(III)Cl3 to 100 mM with ultrapure H2O, and calculate the amount of ultrapure H2O required based on the initial metal weight used in step 1.1 (molecular weight of 58Fe(III)Cl3 is 162.2).
- Leave the ferric chloride solution in the hood at 60 °C with the cap off to evaporate the sample.
- Prepare 58Fe(III)-nitrilotriacetate (NTA) by incubating 58Fe(III)Cl3 with NTA at a 1:5 molar ratio in the presence of 20 mM NaHCO3.
- Prepare 500 mM NTA in 1 N NaOH.
- Prepare 5x transferrin-loading buffer (0.5 M HEPES, pH 7.5; 0.75 M NaCl).
- Prepare 1 M NaHCO3 in ultrapure H2O.
- To a 15 mL conical tube, add 150 µL of 100 mM 58Fe(III)Cl3 solution (from step 1.3.2), 150 µL of 500 mM NTA prepared in 1 N NaOH, 480 µL of ultrapure H2O, 200 µL of 5x transferrin loading buffer, and 20 µL of 1 M NaHCO3 solution.
- Incubate the mixture for 5 min at room temperature.
- Load apo-Tf with 58Fe(III)-NTA to form 58Fe-Tf.
NOTE: This protocol was adapted from McCarthy and Kosman28.- Dissolve 500 mg of apo-Tf in 4 mL of 1x Tf-loading buffer.
- To the 15 mL conical tube in step 1.4.4 containing 1 mL of the 58Fe(III)-NTA solution, add 4 mL of apo-Tf solution.
NOTE: This is a 3:1 molar ratio of 58Fe-NTA with apo-Tf. Each Tf contains 2 Fe binding sites; excess 58Fe-NTA was added to ensure that Tf was fully loaded. - To allow maximal loading of 58Fe-NTA onto apo-Tf, check that the solution is at pH 7.5, and adjust the pH, if necessary, with NaHCO3 or HCl.
- Incubate for 2.5 h at room temperature.
- Remove excess unbound 58Fe(III)-NTA and released NTA.
- Transfer the 58Fe-Tf solution to a molecular weight cutoff column (30 kDa cutoff) and centrifuge at 2,500 × g for 15 min at room temperature.
- Wash the column with 10 mL of 1x transferrin-loading buffer and centrifuge at 2,500 × g for 15 min at room temperature. Repeat the wash and centrifugation, perform a saline wash with 10 mL of saline, and centrifuge at 2,500 × g for 15 min at room temperature.
- Calculate the concentration of 58Fe-Tf.
NOTE: Due to the addition of excess 58Fe in step 1.5.2, assume that all transferrin is diferric. As 500 mg of apo-Tf was used, ~500 mg 58Fe-Tf was produced in step 1.5.4.- Measure the volume recovered from centrifugation after the saline wash in step 1.6.2.
- Divide 500 mg by the volume recovered to determine the concentration (in mg/mL) of the 58Fe-Tf solution.
- Sterilize the 58Fe-Tf solution using a 0.22 µm syringe filter; store at 4 °C until ready to use.
NOTE: 58Fe-Tf solution was used between 1 to 4 weeks post preparation.
2. Set up timed mouse pregnancies
- Use 6- to 8-week-old female mice. Place animals on a low-iron diet (4 ppm iron) or standard chow (185 ppm iron) for 2 weeks prior to mating and maintain animals on the respective diets throughout pregnancy.
- Option 01: Confirm pregnancy by weight gain at E7.5.
- Set up multiple breeding cages. For each cage, combine 2 females with 1 male overnight; the following day when animals are separated is considered embryonic day (E)0.5. Weigh females at E7.5 to determine if pregnant. Mate males again with females that did not gain weight.
NOTE: In WT C57BL/6, a weight gain of 1 g at E7.5 is a good indicator of pregnancy. This method ensures that implantation occurred within a specific 16 h timeframe, allowing for synchronous treatment of all animals that became pregnant during the same mating period.
- Set up multiple breeding cages. For each cage, combine 2 females with 1 male overnight; the following day when animals are separated is considered embryonic day (E)0.5. Weigh females at E7.5 to determine if pregnant. Mate males again with females that did not gain weight.
- Option 02: Confirm pregnancy by plug checks.
- Combine 2 females with 1 male and perform daily plug checks to determine if copulation has occurred.
NOTE: This method may result in staggered pregnancies, and the presence of a plug does not guarantee pregnancy.
- Combine 2 females with 1 male and perform daily plug checks to determine if copulation has occurred.
3. Administer 58Fe-Tf intravenously to E17.5 pregnant mice
- Prepare 58Fe-Tf from step 1.8 for injection.
- Prepare 58Fe-Tf solution at 35 mg/mL in saline; inject 100 µL per mouse.
- Fill an insulin syringe with 100 µL of the 58Fe-Tf solution.
NOTE: Each dose contains 3.5 mg of human 58Fe-Tf (5 µg of 58Fe).
- Anesthetize a pregnant mouse using isoflurane.
- Use an isoflurane regulator with a chamber.
- Use the following settings: 5% isoflurane, 2 L/mL of O2, 2 min.
- Confirm the mouse is anesthetized by looking for lack of response to a toe pinch.
- Apply eye lubricant to the surface of the eye and place the mouse on a heating pad.
- Slowly and carefully inject the 58Fe-Tf solution into the retro-orbital sinus.
- Allow the mouse to recover from anesthesia; do not leave the animal unattended until it has regained sufficient consciousness to maintain sternal recumbency.
- Six hours post injection, euthanize E17.5 pregnant females by isoflurane overdose.
- Perform a cardiac puncture to exsanguinate the mouse as a form of secondary euthanasia.
- Pin the feet down with needles for stabilization.
- Collect the placentae and embryo livers.
- Using sterile forceps and dissection scissors, carefully remove the uterus from the pregnant mouse. Cut off a placental fetal-placental unit, which comprises a single fetus and placenta in the amniotic sac surrounded by a portion of the uterus.
- Carefully cut through the uterus and amniotic sac without disturbing the fetus and placenta.
- Peel back the amniotic sac and remove the fetus and placenta.
- Cut the umbilical cord.
- Blot the fetus and placenta on a clean task wipe to remove the excess amniotic fluid.
- Record the weights of the whole placentae.
- Cut each placenta in half with a razor blade, place each half in a 2.0 mL tube, and snap-freeze in liquid nitrogen.
NOTE: Because 58Fe does not require special handling precautions and disposal, one-half of the placentae can be used for 58Fe measurement and the other half for any other analyses, including quantitation of transferrin receptor (TFR1) and ferroportin (FPN) expression by western blotting and qPCR. - To collect embryo livers, sacrifice the embryo: use a razor blade to rapidly decapitate the embryo.
NOTE: At E17.5, all embryos in the uterus must be euthanized individually, even if they are not used in the study. - Pin down the embryo for stabilization, leaving the abdomen exposed.
- Using dissection scissors, make a small incision where the umbilical cord was attached, insert one end of the dissection scissors into the incision, and perform a median plane cut toward the coronal plane about ¼ inch. Then, perform transverse plane cuts to expose the fetal liver.
- Use forceps to remove the fetal liver.
- Record the weights of the whole embryo livers.
- Place the whole embryo livers in 2 mL tubes and snap-freeze them in liquid nitrogen.
NOTE: Alternatively, only a portion of the embryo liver can be used for 58Fe measurement if additional analyses are desired. Using 2.0 mL tubes allows for better tissue homogenization than 1.5 mL tubes.
- Store the tissues indefinitely at -80 °C.
4. Process tissues for quantitative iron analysis by ICP-MS
- Process the placentae and fetal livers for the quantitation of nonheme iron.
- Thaw placental halves and whole fetal livers, and weigh placental halves (see step 3.6.12 for recording fetal liver weights).
- Add 400 µL of protein precipitation solution (0.53 N HCl, 5.3% TCA).
- Homogenize the tissue using an electric homogenizer.
- Incubate the samples at 100 °C for 1 h.
- Cool the samples in room temperature water for 2 min.
- Open the caps to release pressure, then close the tubes again.
- Centrifuge at 17,000 × g for 10 min at room temperature to pellet tissue debris.
- Carefully transfer the supernatant to a new labeled tube.
- Send samples off for ICP-MS analysis.
- Process the placentae and fetal livers for the quantitation of heme-iron.
NOTE: Following extraction of nonheme iron in step 1, the iron remaining in the pellet is predominantly heme.- Record the weight of each pellet from step 4.1.7.
- Digest the pellets in 10 mL of concentrated 70% HNO3 supplemented with 1 mL of 30% H2O2
NOTE: Consult with the ICP-MS core or center to optimize the volume of HNO3 for specific studies; the volume will partly be dependent on sample weight. - Heat the samples to 200 °C for 15 min.
- Send the samples off for ICP-MS analysis.
NOTE: If distinguishing between heme and nonheme iron sources is not required and only total iron is measured, whole tissue can be digested in HNO3 as the first step.
5. Data analysis
NOTE: Data from ICP-MS has been provided as 56Fe and 58Fe concentrations in ng/mL or mg, ppb (Table 1). 56Fe is the most abundant iron isotope in nature, and its measurement reflects iron accumulation in the placenta/embryo over the entire pregnancy, whereas 58Fe measurement reflects iron that was transferred during 6 h after injection.
- Subtract the natural abundance of 58Fe (0.28% of total Fe) from the measured 58Fe values.
- Calculate total nonheme 58Fe.
- Calculate embryo liver total nonheme iron (ng) by first multiplying the iron concentration (ng/mL) calculated in step 5.1 by the volume (mL) during initial processing in step 4.1.2 to estimate total 58Fe.
- Calculate the amount of iron in the whole placenta by taking the total weight of the placenta measured in step 3.6.6 and dividing it by the weight of the placenta processed in step 4.1.1. Multiply this value by the total nonheme iron (ng) calculated in step 5.2.1 to obtain the total nonheme 58Fe content of the placenta.
- Calculate total heme 58Fe.
- Calculate total heme 58Fe by first multiplying the iron concentration (ng/mg) calculated in step 5.1 by the weight of the pellet (in mg) measured in step 4.2.1.
- Then, divide the total weight of the placenta measured in step 3.5.1 by the weight of the placenta pellet measured in step 4.2.1. Multiply this value by the total heme iron (ng) calculated in step 5.3.1 to obtain total heme 58Fe content of the placenta.
- Sum the calculated nonheme and heme 58Fe values to determine the total iron content for each tissue.
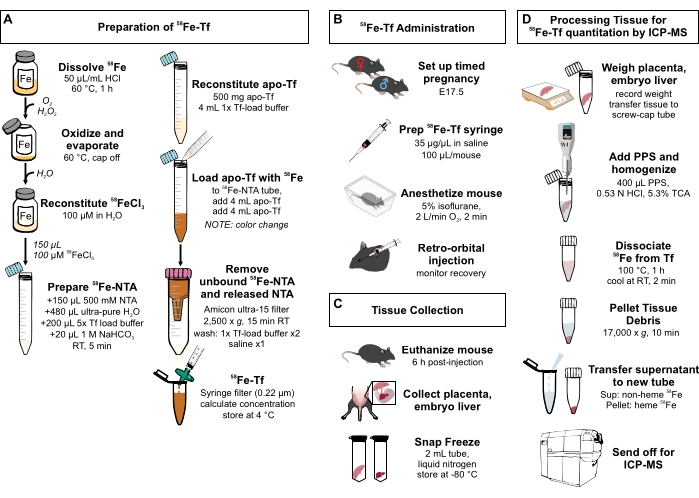
Figure 1: Visual summary of steps in the protocol. (A) Preparation of 58Fe-transferrin. (B) In vivo administration of 58Fe-transferrin. (C) Tissue collection and storage. (D) Processing of the placenta and embryo liver for quantitation of metal species by ICP-MS. Abbreviations: Fe = iron; NTA = nitrilotriacetic acid; Tf = transferrin; PPS = protein precipitation solution; Sup = supernatant; TCA = trichloroacetic acid; ICP-MS = inductively coupled plasma mass spectrometry. Please click here to view a larger version of this figure.
An earlier study using stable iron isotopes to measure iron transport demonstrated that maternal iron deficiency resulted in the downregulation of the placenta iron exporter, FPN4. FPN is the only known mammalian iron exporter, and the absence of FPN during development results in embryonic death before E9.529. To determine whether the observed decrease in FPN expression translated functionally to decreased placental iron transport, 58Fe-Tf was injected intravenously into pregnant dams, and iron in the placenta and the embryo was quantified in the presence of maternal iron deficiency.
To understand how placental iron transport is affected by maternal iron status, iron deficiency was modeled in mice4. Female C57BL/6 mice were placed on a low-iron diet (4 ppm iron) or standard chow (185 ppm iron) for 2 weeks prior to and throughout pregnancy. This dietary regimen results in lower maternal liver nonheme iron and serum iron and hemoglobin at E12.5, E15.5, and E18.5 compared to animals on a standard diet4. At E18.5, embryos from iron-deficient mothers had lower liver iron and were hypoferremic and anemic than embryos from iron-replete mothers. Three pregnant mice were used in each of the iron-replete and iron-deficient groups, and 2-3 placentae were used from each pregnant mouse for analysis.
To quantitate placental iron transport, 58Fe-transferrin was prepared and injected intravenously in pregnant dams and 58Fe measured in the placenta and fetal liver by ICP-MS, as described in the protocol and illustrated in Figure 1. Prior to sending nonheme iron samples out for ICP-MS analysis, total nonheme iron levels were independently quantified via a ferene method described previously30. Nonheme iron concentrations measured by the ferene versus ICP-MS methods were highly significantly correlated in all tissues measured (R2 = 0.94, P < 0.0001, n = 36). Representative results from ICP-MS quantitation of iron isotopes are presented in Table 1. Total 58Fe was calculated as described in step 5 of the protocol. Data are presented as total rather than heme or nonheme iron (Figure 2A-D) because the aim was to quantitate total iron transferred into the placenta and total iron transferred to the embryo from the placenta.
On average, 21% of the administered 58Fe dose was recovered in the placenta, embryo liver, and embryo serum combined. The 56Fe measurement provides insight into the long-term iron transfer in the placenta and embryo liver throughout pregnancy. The total placental 56Fe was similar in the iron-deficient and -replete groups (Figure 2A), whereas the total embryo liver iron was decreased in the iron-deficient group (Figure 2B). This was expected based on the observed decrease in placental FPN in the iron-deficient group4, which would result in iron retention in the placenta at the expense of the embryo. Total 58Fe provides a snapshot of short-term iron transport. In this study, similar to 56Fe, placental 58Fe was similar in both the iron-deficient and -replete groups (Figure 2C), and embryo liver 58Fe was decreased in the iron-deficient group (Figure 2D). These data indicate that during iron-deficient pregnancy, the downregulation of placental FPN results in decreased iron transport to the embryo, leading to cumulative differences in iron content in the placenta and embryo.
It is important to consider the dose of iron administered as it could lead to unintended changes in hepcidin concentration or iron transporter expression31. It was demonstrated that maternal iron deficiency caused a decrease in placental FPN4. To determine if Fe-Tf injection affected this regulation, placenta FPN was measured 6 h post injection by western blot. The iron dose of 5 µg was insufficient to alter placental FPN regulation by maternal iron deficiency (Figure 3).
In summary, this method was used to demonstrate that physiological regulation of placental FPN during maternal iron deficiency results in decreased iron transport across the placenta in vivo. Stable iron isotopes provide a sensitive and quantifiable alternative to radioactivity for the measurement of iron transport and distribution, allowing the simultaneous use of tissues for additional analyses.
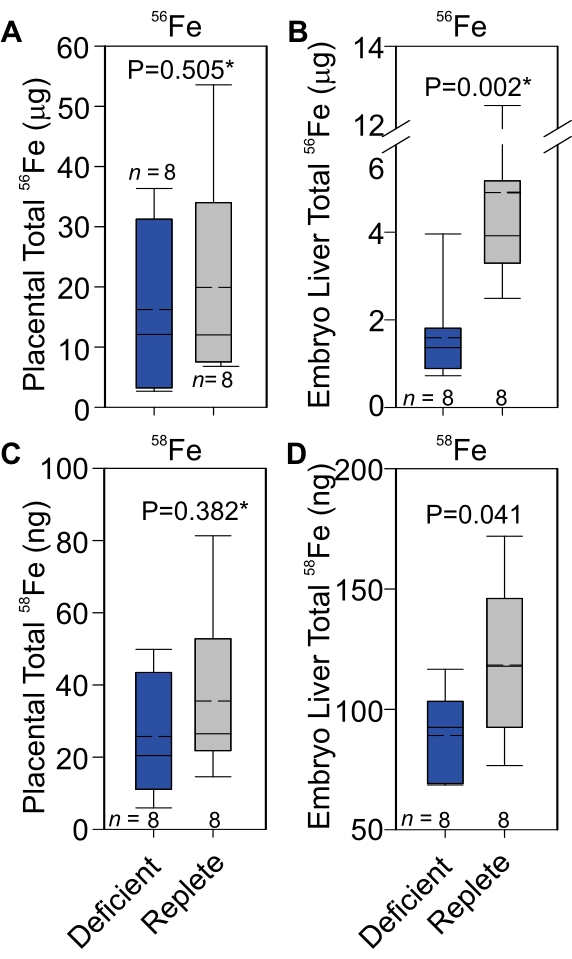
Figure 2: 56Fe and 58Fe transport across the placenta in iron-deficient or iron-replete pregnancies. Total 56Fe in the placenta (A) and embryo liver (B). Total 58Fe in the placenta (C) and fetal liver (D). Statistical analysis was performed using a 2-tailed Student's t-test for normally distributed values and otherwise by Mann-Whitney U rank-sum test (denoted by an asterisk after the P-value). The number of animals is indicated in the x-axes of the box and whisker plots. The upper portion of the box plot indicates the 75th percentile, and the bottom indicates the 25th percentile; whiskers above the box indicate the 90th percentile, and those below the box indicate the 10th percentile. The solid line within the box indicates the median and the dashed line the mean. Statistical analysis was performed using scientific graphing and data analysis software. This figure has been modified from4. Abbreviation: Fe = iron. Please click here to view a larger version of this figure.
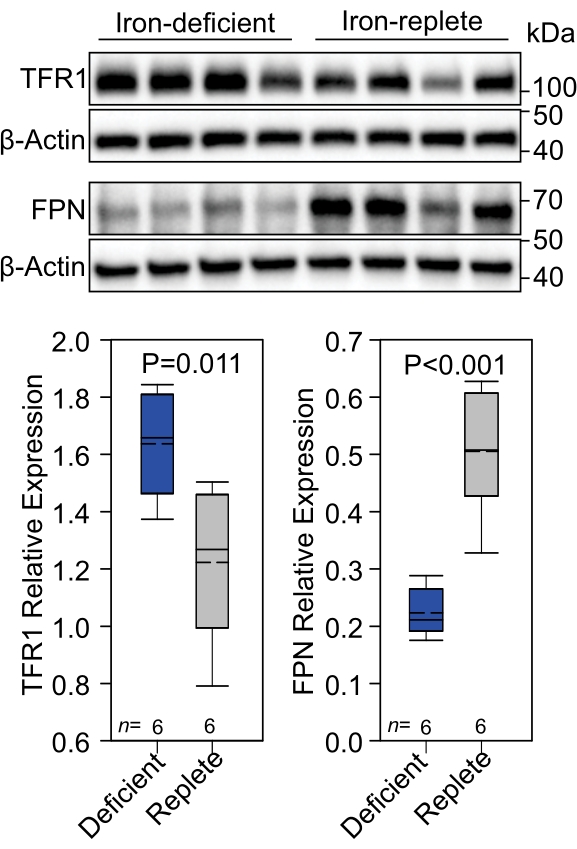
Figure 3: Placental TFR1 and FPN levels. (A) TFR1 and FPN expression was assessed by western blot in iron-deficient and -replete placentae 6 h post treatment of mothers with 58Fe-Tf. (B) Protein expression was quantitated and presented as protein expression relative to β-actin. Statistical analysis was performed using a 2-tailed Student's t-test for normally distributed values. The number of animals is indicated in the x-axes of the box and whisker plots. The upper portion of the box plot indicates the 75th percentile, and the bottom indicates the 25th percentile; whiskers above the box indicate the 90th percentile, and those below the box indicate the 10th percentile. The solid line within the box indicates the median and the dashed line the mean. Statistical analysis was performed using scientific graphing and data analysis software. This figure has been modified from4. Abbreviations: TFR1 = transferrin receptor; FPN = ferroportin. Please click here to view a larger version of this figure.
| Sample | 56Fe | 58Fe | Total Fe | ||||
| Concentration [ng/mL or mg, ppb] | Concentration [ng/mL or mg, ppb] | Sum of Isotopes [ng/mL or mg] | |||||
| Average* | stdev | Average* | stdev | ||||
| Nonheme iron | Placenta | iron-deficient | 729.7 | 17.7 | 2.5 | 0.5 | 732.2 |
| 704.9 | 6.2 | 3.8 | 0.1 | 708.8 | |||
| 649.8 | 3.8 | 0.0 | 0.0 | 649.8 | |||
| 799.2 | 4.6 | 3.8 | 0.2 | 803.0 | |||
| iron-replete | 1919.1 | 5.3 | 11.0 | 0.2 | 1930.1 | ||
| 1610.0 | 26.8 | 11.7 | 0.6 | 1621.7 | |||
| 1925.5 | 39.0 | 14.0 | 0.3 | 1939.5 | |||
| 2551.6 | 16.1 | 8.3 | 0.4 | 2559.9 | |||
| Heme | Placenta | iron-deficient | 253.8 | 1.8 | 1.1 | 0.0 | 254.9 |
| 32.9 | 0.4 | 0.3 | 0.0 | 33.2 | |||
| 337.7 | 5.1 | 1.4 | 0.0 | 339.1 | |||
| 402.3 | 5.3 | 1.7 | 0.0 | 404.0 | |||
| iron-replete | 123.5 | 1.3 | 0.6 | 0.0 | 124.0 | ||
| 75.7 | 1.3 | 0.4 | 0.0 | 76.1 | |||
| 441.9 | 3.0 | 1.9 | 0.0 | 443.8 | |||
| 250.4 | 1.1 | 1.1 | 0.0 | 251.5 | |||
| Nonheme iron | Embryo Liver | iron-deficient | 361.6 | 8.3 | 31.9 | 1.0 | 393.5 |
| 652.4 | 3.4 | 61.7 | 0.3 | 714.1 | |||
| 411.9 | 10.7 | 43.1 | 0.8 | 455.0 | |||
| 631.1 | 7.5 | 62.8 | 0.2 | 693.9 | |||
| iron-replete | 7657.5 | 129.3 | 226.4 | 2.2 | 7883.8 | ||
| 3820.2 | 69.5 | 119.4 | 3.4 | 3939.6 | |||
| 5519.0 | 112.9 | 145.6 | 0.5 | 5664.6 | |||
| 4617.4 | 78.6 | 91.6 | 1.0 | 4709.0 | |||
| Heme | Embryo Liver | iron-deficient | 44.5 | 0.3 | 1.6 | 0.0 | 46.0 |
| 31.0 | 0.4 | 2.9 | 0.0 | 34.0 | |||
| 11.8 | 0.2 | 1.1 | 0.0 | 12.9 | |||
| 42.3 | 0.1 | 3.2 | 0.0 | 45.5 | |||
| iron-replete | 54.3 | 1.4 | 2.1 | 0.0 | 56.4 | ||
| 31.9 | 0.8 | 1.3 | 0.1 | 33.2 | |||
| 59.4 | 0.6 | 2.2 | 0.0 | 61.6 | |||
| 66.7 | 0.6 | 2.1 | 0.0 | 68.8 | |||
Table 1: Representative results from ICP-MS quantitation of 56Fe and 58Fe in placentae and embryo livers. Abbreviations: ppb = parts per billion; stdev = standard deviation; ICP-MS = inductively coupled plasma mass spectrometry.
