After necroptosis induction, RIPK1 and RIPK3 showed almost identical amyloid-like patterns (lanes 2 and 4, Figure 1). However, MLKL fibers were more heterogeneous and seemed to be smaller than RIPK1/RIPK3 fibers (lane 6, Figure 1). That prompted us to develop the 2D SDD-AGE method to address the possibility whether MLKL forms RIPK1/RIPK3-independent fibers or MLKL simply dissociates from the large RIPK1/RIPK3/MLKL fibers during SDD-AGE.
A general schematic of the electrophoresis and transfer procedure is shown in Figure 2. During the first dimension SDD-AGE, amyloid or amyloid-like fibers will exhibit a characteristic smear, as seen in Figure 3A. Upon running the second dimension SDD-AGE, there are two possible outcomes. In the case of no degradation or dissociation during the gel running process, the fibers will migrate identically during the second run as they did in the first run. In this case, a Western blot will exhibit a diagonal pattern on the membrane at a 45° angle as seen in Figure 3B. If the fibers dissociate during the SDD-AGE process, the smaller fibers will migrate faster during the second electrophoresis. In this case, a Western blot will exhibit vertical streaking below the diagonal line, as shown in Figure 3C.
In the case of the MLKL fibers seen during necroptosis, they show the characteristic smear during the first dimension SDD-AGE (Figure 4A). When those same fibers were subjected to 2D SDD-AGE, they exhibited a sharp diagonal line with no vertical streaking (Figure 4B). With this evidence, it was concluded that MLKL was not dissociating from the fibers during the SDD-AGE process and that the MLKL-containing fibers seen in Figure 1 were indeed distinct from the RIPK1/RIPK3 fibers. Furthermore, when RIPK3 was immuno-depleted from the cell lysates, the MLKL fibers remained intact4, confirming that MLKL fibers and RIPK1/RIPK3 fibers are indeed distinct entities.
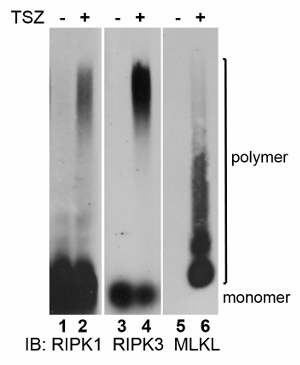
Figure 1. Examination of amyloid-like fibers during necroptosis. Whole cell lysates were harvested from cells undergoing TNF-induced necroptosis and subjected to SDD-AGE. Western blots were performed for RIPK1, RIPK3, and MLKL. The MLKL-containing fibers exhibited a distinct migration pattern from the RIPK1/RIPK3-containing fibers. The MLKL monomer is barely visible under this condition. Please click here to view a larger version of this figure.
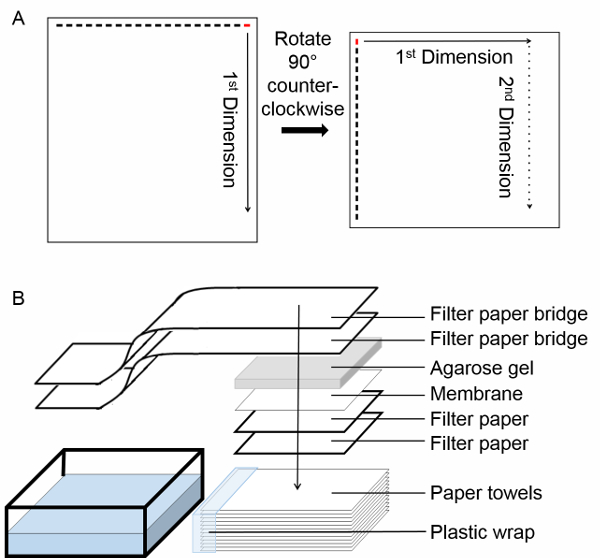
Figure 2. Experimental protocol. (A) Schematic of the two-dimensional semi-denaturing detergent agarose gel electrophoresis (2D SDD-AGE). Begin by loading the sample in the right most lane, labeled in red. After termination of the first dimension run, rotate the gel 90° counter-clockwise. Run the second dimension electrophoresis. Solid arrow indicates the direction of the first dimension run, dotted arrow indicates the direction of the second dimension run. (B) Schema of transfer by capillary action. Please click here to view a larger version of this figure.

Figure 3. Possible results. (A) The expected migration pattern of amyloid or amyloid-like fibers after the first dimension of SDD-AGE. Solid arrow indicates the direction of migration. (B) The expected migration pattern of the SDD-AGE-stable (no degradation or dissociation) amyloid or amyloid-like fibers after 2D SDD-AGE. Solid arrow indicates the direction of the first dimension SDD-AGE. Dotted arrow indicates the direction of the second dimension SDD-AGE. (C) The expected migration pattern of the non-SDD-AGE-stable amyloid or amyloid-like fibers. Solid arrow indicates the direction of the first dimension SDD-AGE. Dotted arrow indicates the direction of the second dimension SDD-AGE. Please click here to view a larger version of this figure.
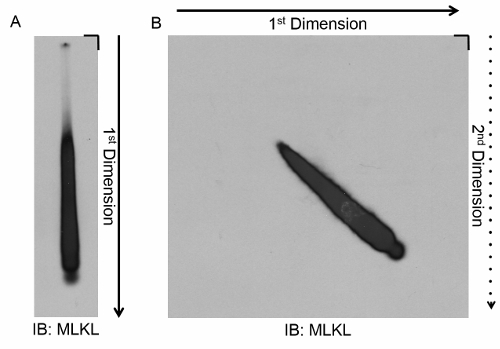
Figure 4. Migration of MLKL-containing fibers during 1D and 2D SDD-AGE. (A) Whole cell lysates were harvested from cells stimulated with TSZ to undergo necroptosis. The lysates were subjected to SDD-AGE, and a Western blot was performed for MLKL. A characteristic smear was observed. (B) Whole cell lysates were harvested from cells stimulated with TSZ to undergo necroptosis. The lysates were subjected to 2D SDD-AGE and a Western blot was performed for MLKL. A sharp line at a 45° angle was observed, indicating that the fibers do not undergo dissociation during SDD-AGE. Please click here to view a larger version of this figure.
