لفهم وظائف الرنا الحفاز وغير الترميز المشاركة في تنظيم الربط والترجمة وتكاثر الفيروس وسرطان، لا بد من معرفة تفصيلية للبنية الحمض النووي الريبي 1،2. للأسف، تنبؤ دقيق من الحمض النووي الريبي للطي يمثل تحديا هائلا. وكلاء التحقيق الكلاسيكية تعاني من عيوب كثيرة مثل سمية، والتغطية النوكليوتيدات غير مكتملة و / أو إنتاجية محدودة إلى 100-150 النيوكليوتيدات في التجربة. خوارزميات التنبؤ المجردة هيكل الثانوي غير ملائم وبالمثل، نظرا لعدم الدقة الناجمة عن عدم قدرتها على التمييز بين فعالية هياكل مماثلة بقوة. الرنا كبيرة على وجه الخصوص هي أيضا في كثير من الأحيان إلى صهر أساليب تحديد هيكل 3D مثل البلورات بالأشعة السينية والرنين المغناطيسي النووي (NMR) التحليل الطيفي، وذلك بسبب مرونتها ومتعلق بتكوين كميات كبيرة من عينات نقية للغاية المطلوبة لهذه التقنيات.
HSHAPE IGH الإنتاجية يحل الكثير من هذه المشاكل عن طريق توفير نهج بسيطة فعالة ليحقق في هياكل الرنا كبير في قرار النوكليوتيدات واحد. وعلاوة على ذلك، والكواشف المستخدمة لشكل هي آمنة وسهلة للتعامل، وعلى النقيض من معظم الكواشف الكيميائية الأخرى التحقيق، تتفاعل مع جميع ribonucleotides أربعة. ويمكن لهذه الكواشف أيضا اختراق الأغشية الخلوية، مما يجعل من الممكن للتحقيق في الرنا في سياق الجسم الحي (ق) 3. وضعت أصلا في الأسابيع مختبر 4، وقد استخدمت SHAPE لتحليل مجموعة متنوعة واسعة من الرنا، والمثال أبرزها تقرير للهيكل الثانوي كاملة من ~ 9 KB HIV-1 الحمض النووي الريبي الجينوم 5. الإنجازات البارزة الأخرى باستخدام SHAPE تشمل توضيح لهياكل أشباه الفيروسات المعدية 6، الرناوات غير الترميز طويلة الإنسان 7، 8 ريبوسوم الخميرة، وriboswitches 9 وكذلك لتحديد مواقع الربط البروتين في الفيريون المرتبطة HIV-1 RNA 3. WHوقد نشرت إيل الاختلافات الأصلي وعالية الإنتاجية من البروتوكول SHAPE في أماكن أخرى 10-12، ويوفر هذا العمل وصفا مفصلا لتحديد الحمض النووي الريبي هيكل الثانوي بحلول SHAPE الإنتاجية العالية باستخدام [أليغنوكليوتيد الفلورسنت، وكولتر CEQ 8000 محلل الوراثية بيكمان، و SHAPEfinder وRNAStructure (V5.3) والبرمجيات. كما تشمل التفاصيل الفنية لم تنشر من قبل والمشورة استكشاف الأخطاء وإصلاحها.
الاختلافات من الشكل
جوهر الشكل وتنوعاتها هو التعرض من الحمض النووي الريبي في محلول مائي لالانهيدريدات بالإلكترونات التي acylate انتقائي 2'-الهيدروكسيل (OH-2') مجموعات الريبوز، وتنتج adducts ضخمة في مواقع التعديل. هذا التفاعل الكيميائي يخدم كوسيلة لاستجواب ديناميات RNA المحلية الهيكلي، كما النيوكليوتيدات الذين تقطعت بهم السبل واحد هم أكثر عرضة للاعتماد التشكل مواتية لهجوم من قبل بالإلكترونات هذه الكواشف، بينما قاعدة الاقتران أو للإنشاء معمارياالنيوكليوتيدات ained أقل أو يتفاعل 10. تم الكشف عن مواقع تشكيل معقد إضافي عن طريق النسخ العكسي بدء الاشعال من fluorescently أو رديولبلد المهجنة إلى موقع معين على الحمض النووي الريبي المحورة (و"(+)" رد فعل الإرشاد التمهيدي). عندما يفشل الناسخ العكسي (RT) لاجتياز ribonucleotides acylated، ويتم إنتاج مجموعة من المنتجات [كدنا] الذي يتزامن مع مواقع من تعديل أطوال. A السيطرة "(-)" التمهيدي التمديد يتم تنفيذ رد فعل باستخدام الحمض النووي الريبي التي لم تتعرض للكاشف أيضا بحيث الإنهاء المبكر من توليف الحمض النووي (أي "توقف") بسبب الحمض النووي الريبي هيكل، غير محدد RNA حبلا الكسر، وغيرها، قد. يمكن تمييزها عن التوقف التي تنتجها التعديل الكيميائي. وأخيرا، يتم استخدام اثنين من ردود الأفعال dideoxy التسلسل بدء من نفس الاشعال كعلامات لربط النيوكليوتيدات رد الفعل مع تسلسل الحمض النووي الريبي الأولية بعد الكهربائي.
في التطبيق الأصلي من حالات العسر الشديدويستخدم E، وهو نفس نهاية 32 ف المسمى التمهيدي ل(+)، (-)، واثنين من التفاعلات التسلسل. يتم تحميل المنتجات من هذه التفاعلات إلى الآبار المجاورة في 5-8٪ هلام بولي أكريلاميد بلاطة، ومجزأة عن طريق تغيير طبيعة بولي أكريلاميد الكهربائي للهلام (PAGE؛ الشكل 1). التحليل الكمي من الصور هلام التي تنتجها SHAPE التقليدية يمكن القيام بها باستخدام SAFA، شبه الآلي برامج التحليل footprinting 13.
في المقابل، شكل عالية الإنتاجية توظف الاشعال fluorescently المسمى والكهربائي الشعري الآلي. على وجه التحديد، لكل منطقة من الحمض النووي الريبي قيد التحقيق، مجموعة من أربعة الاشعال DNA وجود تسلسل مشتركة ولكن مختلفة 5 'يجب توليفها تسميات الفلورسنت أو شراؤها. هذه أليغنوكليوتيد] بشكل مختلف المسمى تخدم لرئيس اثنين من ردود الفعل SHAPE واثنين من ردود الفعل التسلسل، والمنتجات التي يتم تجميع ومجزأة / الكشف عنها من قبل الكهربائي الشعري الآلي (CE). وهرEAS الملف الشخصى التفاعل من 100-150 NT من الحمض النووي الريبي يمكن الحصول عليها من مجموعة من أربعة تفاعلات باستخدام النهج الأصلي، وشكل عالية الإنتاجية يسمح قرار من 300-600 NT من واحدة عينة مجمعة 3. ما يصل إلى 8 مجموعات من ردود الفعل قد تكون مجزأة في وقت واحد، في حين يمكن للما يصل الى 96 عينات أعدت للتجزئة على مدى 12 أشواط متتالية CE (الشكل 2). وعلاوة على ذلك، فإن البرنامج SHAPEfinder، وضعت لمعالجة البيانات الناشئة من CEQ وتحليل وراثية أخرى وتحليلها، هو أكثر الآلي وتتطلب تدخل المستخدم أقل بكثير من SAFA 13 أو حزم هلام التحليل الأخرى.
ظهرت منهجيات عالية الإنتاجية أكثر تقدما في الآونة الأخيرة مثل PARS (تحليل موازية للبنية الحمض النووي الريبي) 14 وفرج تسلسل (جزء التسلسل) 15، والتي تستخدم الإنزيمات بنية محددة بدلا من الكواشف الألكلة بالتزامن مع الجيل القادم التسلسل التقنيات للحصول على المعلومات!ن حول بنية الحمض النووي الريبي. وعلى الرغم من جاذبية هذه التقنيات، العديد من القيود الملازمة لنوكلياز التحقيق لا تزال 16. يمكن التحايل هذه المشاكل في التسلسل SHAPE (SHAPE تسلسل) 17 البروتوكول، حيث سبق التسلسل التالي جيل عن طريق التعديل الكيميائي والنسخ العكسي من الرنا بطريقة مماثلة لتلك التي يؤديها في الشكل التقليدي. في حين أن هذه الأساليب قد يمثلون مستقبل تحديد بنية الحمض النووي الريبي، من المهم أن نتذكر أن التسلسل التالي جيل مكلفة للغاية، ويبقى غير متاحة لكثير من المختبرات.
تحليل بيانات الشكل
يتم تقديم البيانات المنتجة في محلل الوراثية في شكل رحلاني، حيث كثافة مضان من العينة (ق) التي تتدفق من خلال كاشف الشعرية المرسومة ضد فهرس من وقت الهجرة. هذه المؤامرة تأخذ شكل آثار التداخل المقابلة لقناة مضان أربعةق المستخدمة للكشف عن fluorophores مختلفة، وحيث يتكون كل أثر للقمم المقابلة لمنتجات [كدنا] تسلسل أو الفردية. ويتم تصدير البيانات رحلاني من محلل الوراثية كملف نصي المفصول والمستوردة إلى التحول ShapeFinder وتحليل البرمجيات 18.
يستخدم ShapeFinder في البداية لتنفيذ سلسلة من التحولات رياضية على البيانات لضمان أن أوقات الهجرة وأحجام الذروة تعكس بدقة الهويات وكميات من منتجات التفاعل، على التوالي. ثم يتم محاذاة قمم ومتكاملة، ونتائج جدولتها مع تسلسل الحمض النووي الريبي الأولية. يتم الحصول على "لمحة تفاعل" للجزء ذات الصلة من الحمض النووي الريبي من خلال طرح القيم التحكم من (+) القيم المرتبطة مع بعضها RNA النوكليوتيدات، وتطبيع البيانات كما هو موضح أدناه. يتم استيراد هذا الملف إلى RNAstructure (V5.3) البرامج 19،20، والذي يحول فال تفاعل تطبيعUES إلى القيود الزائفة الطاقة التي يتم دمجها في بنية الحمض النووي الريبي الثانوية خوارزمية قابلة للطي. الجمع بين المواد الكيميائية التحقيق وقابلة للطي الخوارزميات في هذه الطريقة بشكل كبير على تحسين دقة التنبؤ بنية مقارنة إما طريقة وحدها 12،21. إخراج RNAstructure (V5.3) يتضمن صورا لأقل الطاقة RNA الهياكل الثانوية مرمزة مع ملف SHAPE تفاعل (ق)، وكذلك نفس الهياكل في تدوين النصوص الدوت قوس. قد في وقت لاحق يتم تصديرها هذا الأخير إلى برامج مخصصة لعرض رسومية من بنية الحمض النووي الريبي الثانوية مثل فارنا 22 و 23 PseudoViewer.
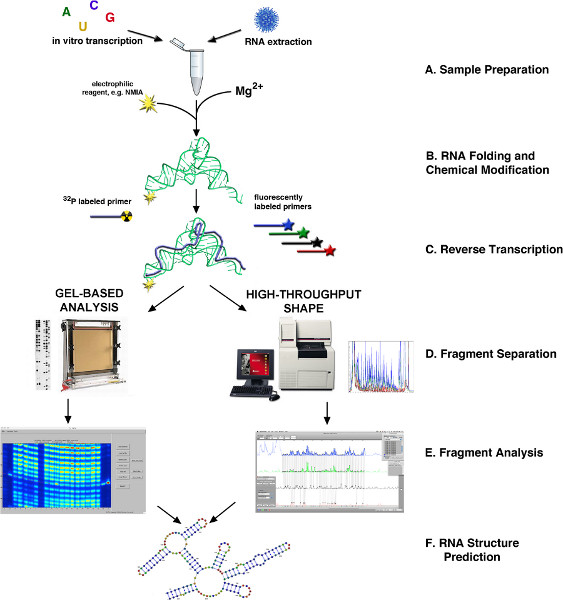
الشكل 1. انسيابي من عزم هيكل الحمض النووي الريبي عبر SHAPE (4،10). (A) RNA ميمكن الحصول على AY من العينات البيولوجية أو بواسطة نسخ في المختبر. (B) اعتمادا على مصدر، يتم طي RNA أو محضرة بطريقة أخرى وتعديلها مع كاشف الشكل. (C) النسخ العكسي باستخدام بادئات fluorescently المسمى أو بالإشعاع. (D) منتجات كدنا] هي مجزأة إما عن طريق الكهربائي الشعري أو لوح المستندة إلى هلام. (E) تحليل شظية. (F) التنبؤ بنية الحمض النووي الريبي. انقر هنا لعرض أكبر شخصية.
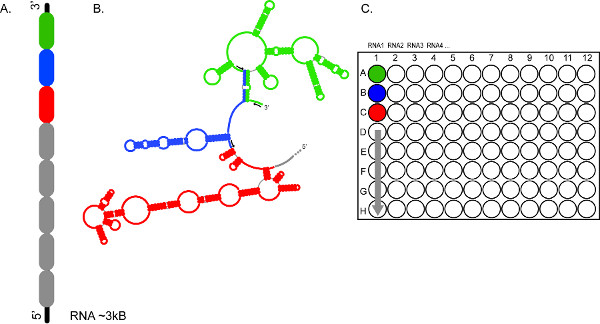
الشكل 2. الطابع الإنتاجية العالية من الشكل القائم على CE يسمح تحليل سريع من الرنا متعددة، و / أو شرائح متعددة من نفس الحمض النووي الريبي. (A) </stرونغ> يمثل كيف يمكن تقسيم على الحمض النووي الريبي إلى 300-600 أقسام NT (ونا مميزا باللون الأخضر والأزرق والأحمر) (B) يتم بحثها أقسام من الحمض النووي الريبي بشكل مستقل باستخدام مجموعات مختلفة من بادئات الفلورسنت (الأسهم السوداء) (C) مجموعة من يتم تجميع ردود الفعل وتحميلها في الآبار A1، B1، C1، الخ، على التوالي، وتوفير تغطية كاملة لل~ 3 KB RNA1. منتجات التفاعل من الرنا 2، 3، 4، وما إلى ذلك قد تكون مستعدة بالمثل لتجزيء في أشواط متتالية الكهربي. انقر هنا لعرض أكبر شخصية.
