A representative example of the application of the microinjection protocol described can be found in Carballar-Lejarazú et al5. The intent here was to insert an autonomous gene-drive system into the germline of a laboratory strain, G3, of An. gambiae. The system was designed to target the cardinal ortholog locus (Agcd) on the third chromosome in this species, which encodes a heme peroxidase that catalyzes the conversion of 3-hydroxykynurenine to xanthommatin, the last step in ommochrome biosynthesis (Figure 11). Homozygous gene drive-containing mosquitoes are loss-of-function cardinal mutants with larvae, pupae and newly-emerged adults having a red-eye phenotype. A plasmid, pCO37, containing the drive system expresses Streptococcus pyogenes Cas9 (SpCas9) endonuclease under control of the regulatory elements of the An. gambiae nanos gene ortholog, a 23-nucleotide guide RNA (gRNA) under the control of an An. gambiae U6 gene promoter and a 3XP3-CFP dominant marker cassette expressing the cyan fluorescent protein for identifying transgenic progeny. Agcd DNA fragments homologous to genomic regions flanking the gRNA target cut site make possible homology-directed repair- (HDR) mediated integration.
Seven-hundred eighty wild-type An. gambiae embryos were injected with the pCO37 plasmid along with SpCas9 protein. One-hundred forty-six injected embryos (18.7%; G0) survived to the adult stage and were outcrossed subsequently to wild-type members of the opposite sex. Fluorescent microscope screening of the next generation (G1) progeny identified three of 3,428 (0.09%) that were positive for the CFP marker gene, only two of which, a male and female, survived. Molecular approaches (gene amplification, DNA sequencing) verified the accurate insertion of ~10 kb of DNA and the resulting strain was designated AgNosCd-1. Extensive follow-up analyses showed that AgNosCd-1 was highly-efficient at drive, created few resistant alleles and had no major genetic load (fitness costs) as a result of the integration and expression of the gene-drive system5. The pCO37 plasmid serves as backbone for developing population modification strains of An. gambiae to prevent malaria parasite transmission.

Figure 1: Egg collection device. The egg collection device is composed of filter paper, nylon mesh, dental film and a retrofitted 50 mL conical tube. The device allows mosquitoes to be deposited by aspirator securely into the container via the slit in dental film. The filter paper and nylon mesh allow water to be absorbed, which provides a damp, but not flooded surface, for oviposition. Please click here to view a larger version of this figure.
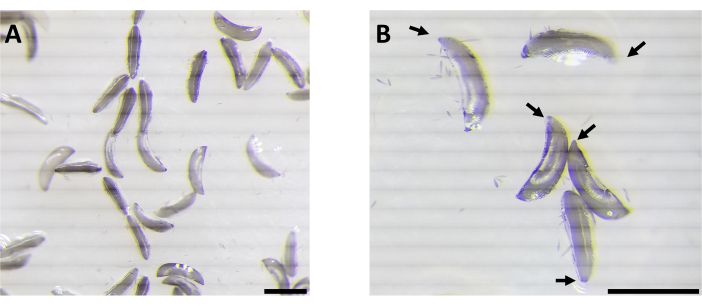
Figure 2: Anopheles gambiae embryos. A) A batch of embryos at varying stages of maturation. B) The arrows indicate the posterior pole, the location within the embryo that contains germline cells, which are the intended targets for transformation. The scale bar represents ~1 mm. Please click here to view a larger version of this figure.

Figure 3: Egg collection. Once the mosquitoes are placed inside the egg collection device, it is moved to a Petri dish containing a layer of filter paper moistened with ddH2O. Adjust the water volume so that the water level rises to ~1/4 the height of the cap. The egg collection device and Petri dish are then placed in a darkened incubator and mosquitoes allowed to oviposit for a total of 45 min. Please click here to view a larger version of this figure.

Figure 4: Embryo development. The embryos are only suitable for injection during a short time window. Eggs that are injected prematurely in development are too delicate and will suffer damage and not hatch. Eggs that are injected later in their development become hardened and can break the needle causing too much genetic material to be injected in the egg. Additionally, injections later in development are less likely to cause permanent changes to the germline as pole cells rapidly undergo further replication and differentiation. Pictures were taken every 10 min after oviposition time of 45 min. The asterisks (*) indicate eggs that are suitable for injection. Please click here to view a larger version of this figure.

Figure 5: Membrane slide assembling. The eggs are aligned along the nylon membrane, which provides a support structure so that the eggs do not move during the injection process. The filter paper serves as a reservoir for water to ensure that the eggs stay moist throughout the injection. Place the filter paper and nylon membrane close to the edge of the glass slide allowing room for both eggs and the water that submerges them. If the membrane and filter paper are too far from the edge of the slide, achieving the correct angle between embryos and needle will be difficult. Please click here to view a larger version of this figure.

Figure 6: Embryo orientation for microinjection. A) Embryos are aligned with the posterior poles lower than the anterior, running down the membrane. B) The embryos are immersed in water; adjust the width of the water line above the egg to be approximately an eighth to a quarter the width of an average embryo. C) Close-up image of needle angle with embryos. The scale bar represents ~1 mm. Please click here to view a larger version of this figure.

Figure 7: Abnormal embryos. Embryos that appear abnormal in coloration or texture will not hatch; do not inject them. Embryos that are yellow in color will not properly melanize. Reduced hatching has also been observed in smooth embryos that do not have normal-appearing eggshells (cuticles). Please click here to view a larger version of this figure.

Figure 8: Needle position during the microinjections. A) Wide-angle view of the injection stage. B,C) Position the needle so that the aligned embryos form an angle of 15° with the injection needle. Do not elevate the arm of the microinjection instrument that holds the needle significantly higher than the microscope stage to ensure proper angling between the needle and slide containing the embryos. Please click here to view a larger version of this figure.

Figure 9: Co-axial controls and needle holder. A) The co-axial controls (arrow) allow for 3-dimensional movement of the needle to ensure precise placement of the needle for each injection. It is important to use co-axial controls to raise the needle vertically when switching slides so that the needle does not collide with the new slide when putting it into place on the stage. Lateral co-axial controls allow for accurate penetration of the needle into the posterior pole. B) Image of needle holder showing angle for injection. Please click here to view a larger version of this figure.

Figure 10: Chamber for egg hatching. A,B) Carefully wash injected embryos into a cylindrical container filled to a quarter depth with double-distilled water and lined with filter paper. C,D) Movement of the container causes the eggs to naturally adhere to the filter paper. Before leaving the container for hatching, ensure any eggs that have adhered higher up the filter paper are gently washed back down to water level. The blue oval marks a number of deposited eggs to show relative size. Please click here to view a larger version of this figure.
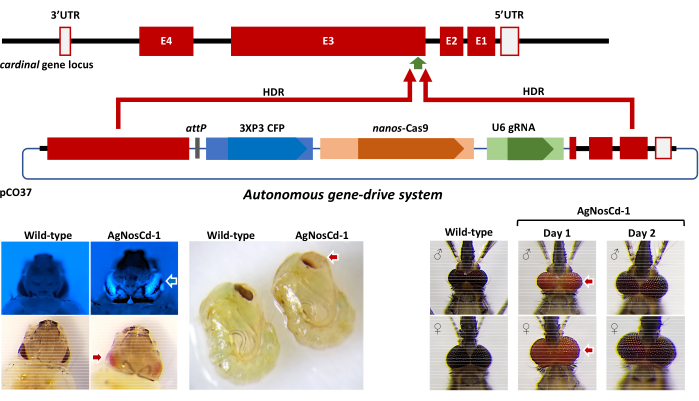
Figure 11. Anopheles gambiae cardinal (Agcd) gene ortholog, pCO37 gene drive construct and resulting phenotypes. Top) Agcd gene: maroon blocks, exons (E1-4); empty blocks, 3'- and 5-untranslated regions (UTR); thick black line, introns and intergenic DNA. Middle) pCO37 plasmid: homology arms from Agcd gene: maroon blocks, blue blocks, dominant marker gene components (3XP3 and CFP); tan blocks, drive components (nanos promoter and SpCas9 protein-encoding sequences); green blocks, guide RNA components (U6 promoter and gRNA sequence). Genes and features of pCO37 are not to scale. Recombination (maroon arrows) resulting from homology directed repair (HDR) initiated at the SpCaS9/gRNA-mediated cut site (green arrowhead) results in integration of the gene drive construct. Bottom) resulting visible phenotypes of gene drive integration: (left) CFP+ (white/blue arrow) and homozygous Agcd-mutant (red arrow) phenotypes in larvae and Agcd-mutant (red arrow) phenotypes in pupae. (right) Homozygous Agcd mutant phenotype "red eye" in adults. Adapted from Carballar-Lejarazú et al. (2020) and used with permission5. Please click here to view a larger version of this figure.
