Our protocol describes the generation of mouse EHBD organoids that are tissue-specific and adult stem cell-derived. After the organoids are cultured, a cystic structure formation can be observed as early as 1 day after the EHBD isolation. Contamination with fibroblasts is not typically observed during culture generation. EHBDO plating efficiency is approximately 2% when isolated from either neonatal or adult (older than 2 months) mice (Figure 2B). Plating efficiency of EHBD organoids derived from adult mice increases to 11% in passage 2 and remains stable (Figure 2B). The majority of organoids demonstrate cystic morphology through all passages, with rare "irregular" organoids (Figure 2C-E). Organoids reach a growth peak at 5-7 days after which they start accumulating intraluminal debris and deteriorate (Figure 2A). Therefore, for maintenance of organoid culture, they should be split every 7-10 days (Figure 2A). Once established and when appropriately handled, organoids can be maintained in culture almost indefinitely (cultures were observed up to 14 months). To avoid culture contamination with differentiated cells carried over from initial cell isolation, use organoids passaged at least twice prior to using them for a downstream application. For long-term storage, use earlier passage (up to passage 7) organoids, since they have higher plating efficiency after recovery from storage.
When analyzed with immunofluorescence, EHBDOs consist of a pure population of epithelial cells marked by E-cadherin (Figure 3A-C). Organoid cells demonstrate markers of biliary progenitor cells (Pancreatic and Duodenal Homeobox 1 (PDX1); Figure 3A) as well as markers of biliary differentiation (cytokeratin 19 (CK19) and Sex-Determining Region Y-Box 9 (SOX9); Figure 3B, C). Importantly, a high percentage of organoid cells possess a primary cilium marked by acetylated α-tubulin (a-AT; Figure 3D), which is a feature of normal cholangiocytes, and suggests appropriate organoid cell polarization. The expression of markers of progenitor (Pdx1) and biliary differentiated cells [Ck19, Sox9, Aquaporin 1 (Aqp1), Cystic Fibrosis Transmembrane Conductance Regulator(Cftr)] can be also confirmed by real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR) (Table 1). Combination of these markers is characteristic for cholangiocytes in EHBDs14,17,18.
In summary, this protocol describes the generation of an organoid culture model of polarized biliary epithelial cells expressing progenitor and differentiated markers. This system can be maintained in culture for a prolonged time without changes in morphology, stored long-term, and analyzed with immunohistochemistry and qRT-PCR.
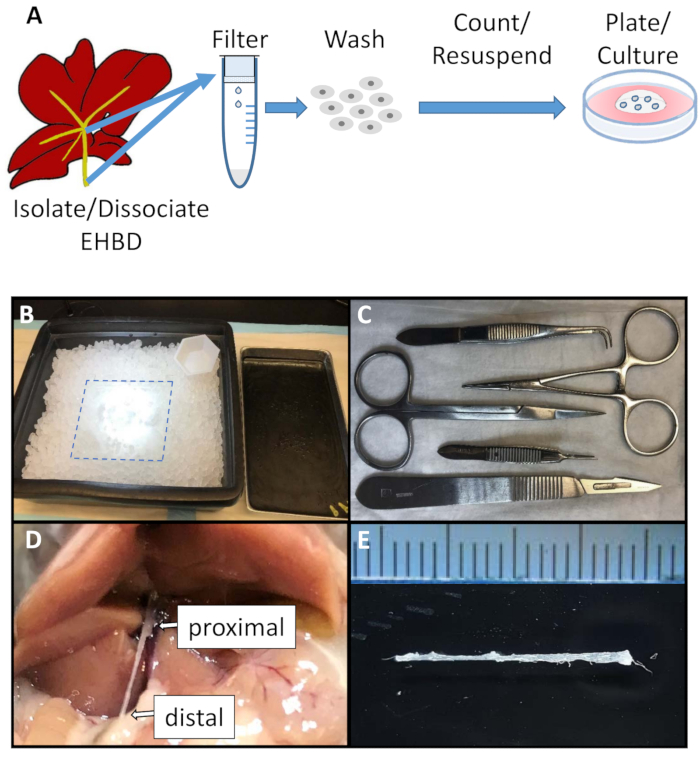
Figure 1: Schematic of the EHBD organoid culture generation and surgical set up. (A). Schematic of EHBD organoid generation. (B). Surgical area was set up for EHBD isolation and included a glass plate (dotted line) kept on an ice tray at all times. (C). Sterile surgical equipment included sharp scissors, straight and curved serrated tweezers, hemostat, and scalpel. (D and E) EHBD is isolated from surrounding connective and pancreatic tissue followed by careful dissection proximally from the intrahepatic bile ducts and liver (D, arrow), and distally from the duodenum (D, arrow). Ruler marks = 1 mm. Please click here to view a larger version of this figure.
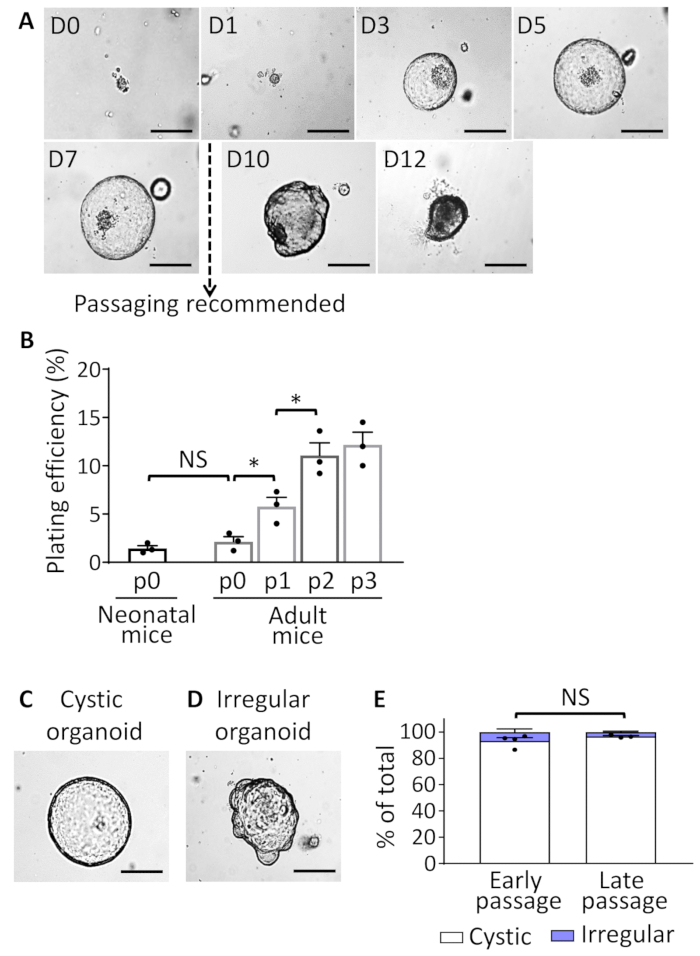
Figure 2: EHBDO culture. (A). Microscopic images of EHBDOs over a 12-day course. (B). Plating efficiency of organoids derived from the neonatal (2 mice per culture, n = 3 cultures) and adult (>2 months old, 1 mouse per culture, n = 3 cultures) mice after plating 300 cells per well in 24-well plate and enumerating established organoids on day 5 of culture. (C and D) EHBDO cystic versus irregular morphology was analyzed by microscopy. (E). The percent of cystic and irregular shaped organoids was analyzed in early (<10) and late (≥10) organoid passages. Scale bars = 500 µm. Quantitative data showed as mean +/- standard error of the mean (SEM), t-test. NS = not significant. Please click here to view a larger version of this figure.
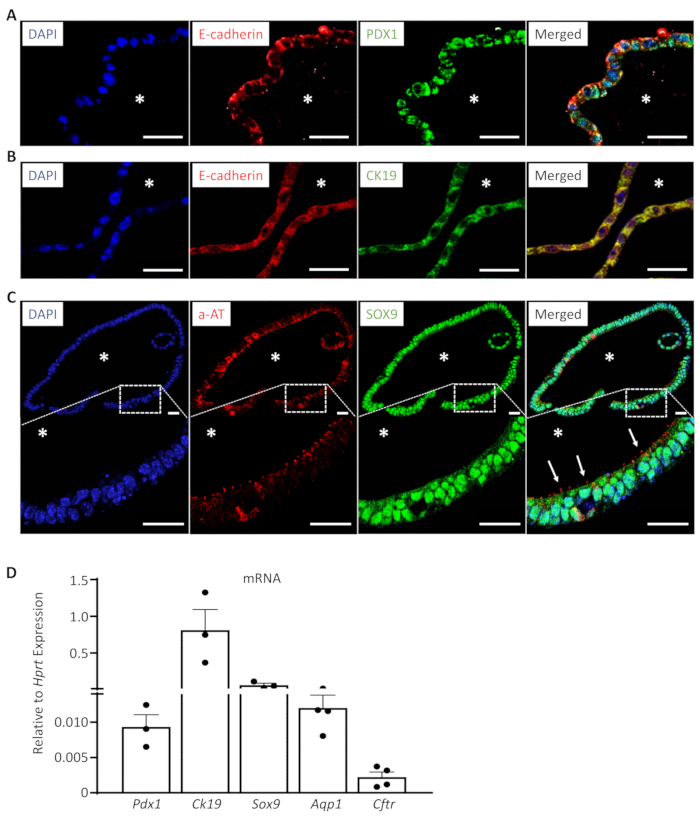
Figure 3: EHBDOs express markers of progenitor and mature biliary cells. (A-C). EHBDOs were analyzed by immunofluorescence staining for markers epithelial (A, B. E-cadherin, red), progenitor (A. PDX1, green), and differentiated (B. CK19, green; and C. a-AT, red) biliary cells. Scale bars = 25 µm. *, lumen. (D). EHBDOs were analyzed for abundance of Pdx1, Ck19, Sox9, Aqp1, and Cftr mRNA by qRT-PCR (mean +/- SEM relative to expression of Hprt). Please click here to view a larger version of this figure.
| Gene | Accession number | Primer sequence | Product size | ||
| Hprt | NM_013556 | Forward 5’-AACTTGCGCTCATCTTAGGCTTTG-3’ | 173 bp | ||
| Reverse 5’-AGGACCTCTCGAAGTGTTGGATAC-3’ | |||||
| Pdx1 | NM_008814 | Forward 5'-GAATTCCTTCTCCAGCTCCA-3' | 133 bp | ||
| Reverse 5'-GATGAAATCCACCAAAGCTCA-3' | |||||
| Sox9 | NM_011448 | Forward 5’-TCCACGAAGGGTCTCTTCTC-3’ | 107 bp | ||
| Reverse 5’-AGGAAGCTGGCAGACCAGTA-3’ | |||||
| Ck19 | NM_008471 | Forward 5’-TCTGAAGTCATCTGCAGCCA-3’ | 133 bp | ||
| Reverse 5’-ACCCTCCCGAGATTACAACC-3’ | |||||
| Aqp1 | NM_007472 | Forward 5’-CAGTACCAGCTGCAGAGTGC-3’ | 112 bp | ||
| Reverse 5’-CATCACCTCCTCCCTAGTCG-3’ | |||||
Table 1: Primers.
