Characterization of Differentiated hiPSC-CMs, -ECs, and -SMCs
The differential capacity of hiPSCs were evaluated2,3,4. Flow cytometry analyses of cardiac troponin T (cTnT) expression suggest that the purity of the final hiPSC-CM population can exceed 90% (Figure 1A, 1B, panel B1). Nearly all of the cells expressed slow myosin heavy chain (Figure 1B, panel A1), α-sarcomeric actin (Figure 1B, panel A2), while approximately 25% expressed the 2v isoform of myosin light chain (MLC2v) (Figure 1B, panel B2), which was found only in ventricular CMs4. The efficiency of the hiPSC-EC differentiation protocol (i.e., the percentage of CD31+ cells) was substantially higher when performed with hiPSCs from the GRiPS line (45.6%, Figure 1C, panel C2) than with PCBC16iPS cells (31.3%, Figure 1C, panel D2); populations of >95% purity were achieved via selection for CD31 expression, and >90% of the selected cells continued to express CD31 or CD144 for up to 4 weeks when cultured in EGM2-MV medium (without FBS) supplemented with B27, VEGF, and SB431542 (Figure 1D)3. More than 94% of the purified hiPSC-SMCs expressed smooth muscle actin (SMA), but synthetic hiPSC-SMCs were more likely than contractile hiPSC-SMCs to express collagen 1, connexin 43, or vimentin (Figure 1E). The migration and proliferation capability of hiPSC-SMCs were evaluated4. Synthetic hiPSC-SMCs were also more migratory and proliferative than contractile hiPSC-SMCs (Figure 1F, panels i and ii), while contractile SMCs contracted more strongly in response to carbachol treatment (Figure 1F, panels iii, iv and v).
Growth-factor Release from Gelatin Microspheres
ELISA measurements of IGF-1 levels in the medium from cultured, cell-free patches indicated that the growth factor was released from the microspheres over a period of at least 3 days (Figure 2A)2. The analyses were performed by loading 5 mg of IGF-containing microspheres with 5 µg IGF-1. A patch was generated by mixing the microspheres with 1 ml fibrinogen solution and 1 ml thrombin. The patch was cultured in 2 ml MEM. One ml of the medium was collected and replaced with 1 ml of fresh MEM each day (Figure 2B). Data were presented as Mean ± SEM.
Observations from an IR-injury Model
The effectiveness of our patch-mediated cell-transplantation protocol has been investigated in a swine model of ischemia-reperfusion injury2. A total of 6 million hiPSC-CMs, -ECs, and -SMCs (2 million of each cell type) were injected directly into the injured myocardial tissue. For animals in the CELL+Patch group, a patch composed of fibrin and IGF-1-containing microspheres was created over the site of injury before injection, while animals in the CELL group were treated with the same cell dose but without the patch; both treatments were withheld from animals in the MI group. Four weeks after injury and treatment, 9% of the cells delivered to animals in the CELL+Patch group were retained and continued to survive at the site of administration, compared to just 4% of the cells administered to animals in the CELL group (Figure 3A). Treatment with both the cells and the patch, but not with the cells alone, was also associated with significant improvements in measurements of cardiac function and infarct size (Figure 3B).
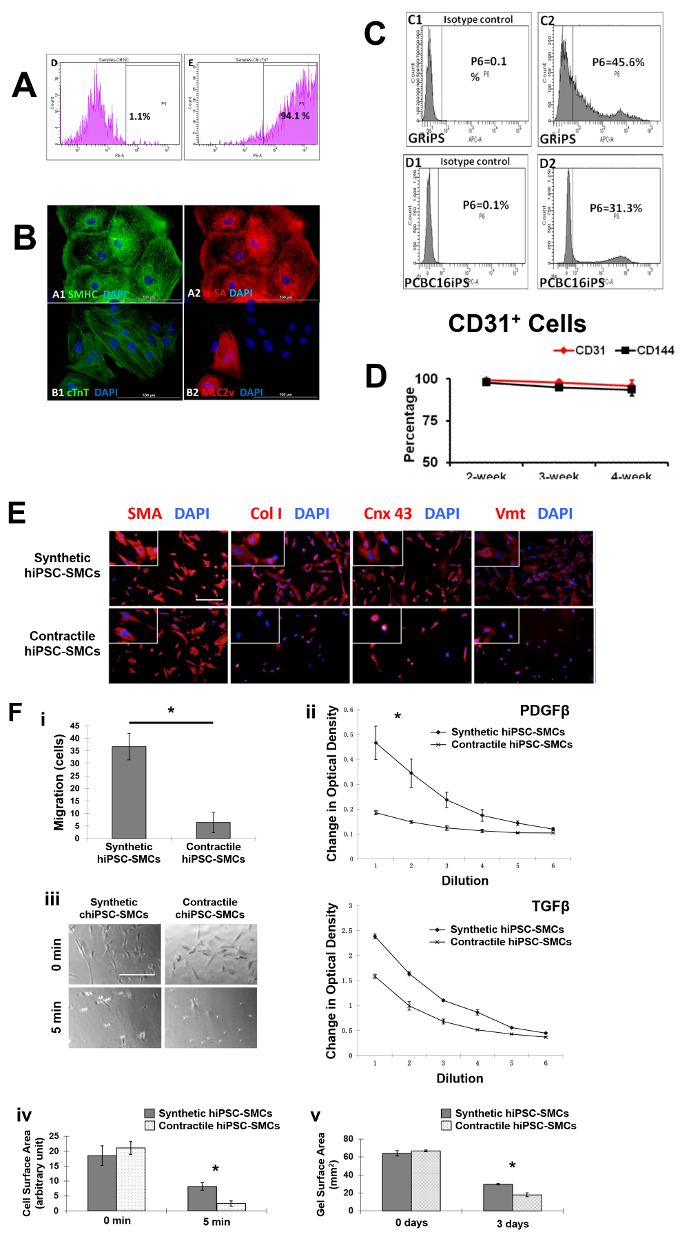
Figure 1: Differentiation of hiPSCs into cardiomyocytes, endothelial cells, and smooth muscle cells. Cardiomyocyte differentiation and the purity of hiPSC-CMs were evaluated via flow cytometry (A) and histology with different cardiac markers (B). The endothelial differentiation of hiPSC was determined with flow cytometry (C). The maintenance of the endothelial phenotype of hiPSC-ECs was assessed via analyzing the expression of CD31 and CD144 at 2, 3, and 4 weeks after the purification by flow cytometry (D). The smooth muscle cell differentiation of hiPSC was measured by various markers (E). And the migration and proliferation potential of synthetic smooth muscle cells vs. contractile smooth muscle cells derived from hiPSCs was assessed (F). Nuclei were counterstained with DAPI in the immunofluorescence staining. Scale bar = 100 µm in (B) and 200 µm (E and F). The migration and proliferation capability of synthetic hiPSC-SMCs were evaluated (F, panel i and ii). And contraction of contractile SMCs in response to carbachol treatment was assessed (F, panel iii-v). *p < 0.05, synthetic hiPSC-SMCs vs. contractile hiPSC-SMCs. A and B were modified from Ye L, et al.2. C and D were modified from Zhang S, et al.3. E and F were modified from Yang L, et al.4. Data were presented as mean ± standard error of the mean (SEM). Comparison between two groups was evaluated via the Student's T test, and comparison between multiple groups was assessed via one-way ANOVA. p <0.05 was considered significant. Please click here to view a larger version of this figure.
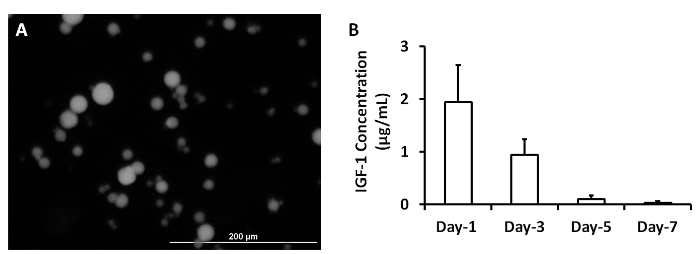
Figure 2: Release of growth factors from microspheres. IGF-containing microspheres was synthesized (A). IGF-1 release from microspheres was measured at 1, 3, 5 and 7 days after they were synthesized (B). Scale bar = 200 µm. Panel A was modified from Ye L., et al.2. Please click here to view a larger version of this figure.
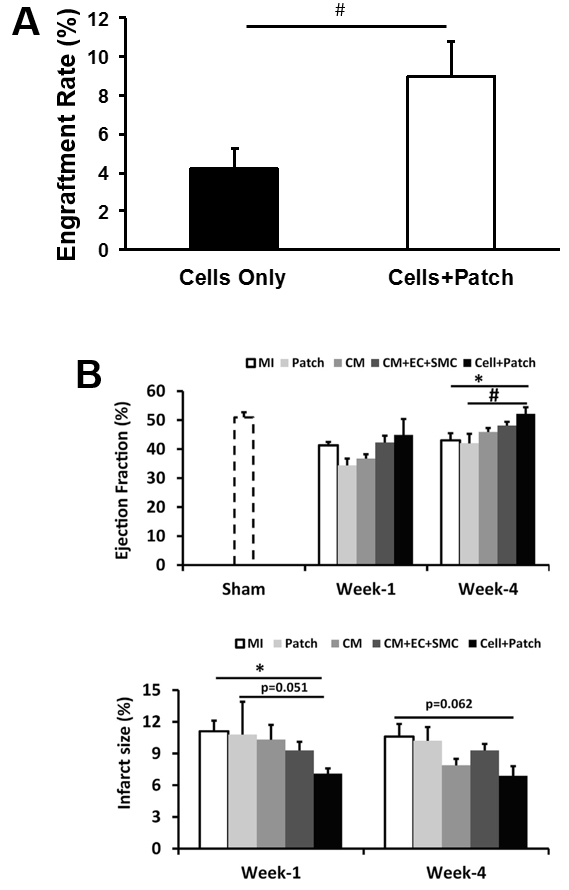
Figure 3: Assessment of efficiency of cell therapy with hiPSC-derived trilineage cells. The overall engraftment rate for the three cell population was assessed 4 weeks after cell transplantation (A). Cardiac function (as indicated by ejection fraction) and infarction size were evaluated via cardiac MRI at 1 and 4 weeks after cell transplantation (B). *p <0.05 versus MI; #p <0.05 versus Patch. A and B were modified from Ye L., et al.2. Data were presented as mean ± SEM. Comparisons among groups were analyzed for significance with one-way ANOVA. p <0.05 was considered significant. Results identified as significant via ANOVA were reanalyzed with the Tukey correction.
| Protocol 1 | |||
| mTeSR1 medium | Stem cell technologies | 5850 | |
| Growth-factor-reduced matrigel | Corning lifescience | 356231 | |
| Y-27632 | Stem cell technologies | 72304 | |
| B27 supplement, serum free | Fisher Scientific | 17504044 | |
| RPMI1640 | Fisher Scientific | 11875-119 | |
| Activin A | R&D | 338-AC-010 | |
| BMP-4 | R&D | 314-BP-010 | |
| bFGF | R&D | 232-FA-025 | |
| Collagenase IV | Fisher Scientific | NC0217889 | |
| Hanks Balanced Salt Solution (Dextrose, KCl, KH2PO4, NaHCO3, NaCl, Na2HPO4 anhydrous) | Fisher Scientific | 14175079 | |
| Fetal Bovine Serum | Fisher Scientific | 10438018 | |
| 6-well plate | Corning Lifescience | 356721 | |
| 10cm dish | Corning Lifescience | 354732 | |
| Cell incubator | Panasonic | MCO-18AC | |
| Materials | Company | Catalog Number | Comments |
| Protocol 2 | |||
| Versene | Fisher Scientific | 15040066 | |
| Fibrinogen | Sigma-Aldrich | F8630-5g | |
| Thrombin | Sigma-Aldrich | T7009-1KU | |
| EMB2 medium | Lonza | CC-3156 | |
| VEGF | ProSpec-Tany | CYT-241 | |
| EPO | Life Technologies | PHC9431 | |
| TGF-ß | Peprotech | 100-21C | |
| EGM2-MV medium | Lonza | CC-4147 | |
| SB-431542 | Selleckchem | S1067 | |
| CD31 | BD Bioscience | BDB555445 | |
| CD144 | BD Bioscience | 560411 | |
| 15 mL centrifuge tube | Fisher Scientific | 12565269 | |
| Eppendorff Centrifuge | Eppendorf | 5702R | |
| Materials | Company | Catalog Number | Comments |
| Protocol 3 | |||
| CHIR99021 | Stem cell technologies | 720542 | |
| PDGF-ß | Prospec | CYT-501-10ug | |
| Materials | Company | Catalog Number | Comments |
| Protocol 4 | |||
| Olive oil | Sigma-Aldrich | O1514 | |
| Gelatin | Sigma-Aldrich | G9391 | |
| Acetone | Sigma-Aldrich | 179124 | |
| Ethanol | Fisher Scientific | BP2818100 | |
| Glutaraldehyde | Sigma-Aldrich | G5882 | |
| Glycine | Sigma-Aldrich | G8898 | |
| IGF | R&D | 291-G1-01M | |
| Bovine serum albumin | Fisher Scientific | 15561020 | |
| Heating plate | Fisher Scientific | SP88850200 | |
| Water bath | Fisher Scientific | 15-462-10Q | |
| Materials | Company | Catalog Number | Comments |
| Protocol 5 | |||
| CaCl2 | Sigma-Aldrich | 223506 | |
 -aminocaproic acid -aminocaproic acid |
Sigma-Aldrich | A0420000 | |
| MEM medium | Fisher Scientific | 12561-056 | |
| Syringe | Fisher Scientific | 1482748 | |
| Anesthesia ventilator | Datex-Ohmeda | 47810 | |
| Anesthesia ventilator | Ohio Medical | V5A | |
| Defibrillator | Physiol Control | LIFEPAK 15 | |
| 1.5T MRI | General Electric | Signa Horizon LX | |
| 7T MRI | Siemens | 10018532 | |
| Gadolinium Contrast Medium (Magnevist) | Berlex | 50419-188-02 | |
| 2-0 silk suture | Ethilon | 685H | |
| 3-0 silk suture | Ethilon | 622H | |
| 3-0 monofilament suture | Ethilon | 627H |
