Das zweigliedrige GAL4-UAS-System ist das Arbeitspferd der funktionellen Charakterisierung von Genen im Insektenmodellorganismus Drosophila melanogaster1,2,3. Um das GAL4-UAS-System zu verwenden, werden transgene Treiberlinien, die den Hefetranskriptionsfaktor GAL4 unter Kontrolle einer regulatorischen Sequenz exprimieren, mit Responderlinien gekreuzt, die ein Gen von Interesse oder ein RNAi-Konstrukt (RNA Interference) tragen, das von einer von GAL4 erkannten Upstream Activation Sequence (UAS) gesteuert wird. Die Nachkommen dieses Kreuzes drücken das interessierende Transgen in einem raumzeitlichen Muster aus, das vom Promotor diktiert wird, der die GAL4-Expression kontrolliert (Abbildung 1). Phänotypen, die von Nachkommen von Driver-Responder-Kreuzungen gezeigt werden, können bewertet werden, um die Funktion von Kandidatengenen aufzuklären. Obwohl D. melanogaster verwendet wurde, um Gene aus anderen Organismen zu untersuchen4,5,6,7, wurde das GAL4-UAS-System nun für den Einsatz bei Insekten von medizinischer und landwirtschaftlicher Bedeutung angepasst, um eine direkte Analyse der interessierenden Arten zu ermöglichen 8,9,10,11,12,13,14.
Bei der afrikanischen Malariamücke Anopheles gambiae wurde das GAL4-UAS-System zunächst mittels Zelllinien-Co-Transfektion9 getestet. Mehrere Konstrukte wurden auf Effizienz in verschiedenen paarweisen Kombinationen untersucht und fanden heraus, dass 14 tandemly wiederholte UAS, ergänzt mit einem kleinen künstlichen Intron (UAS-14i), das breiteste Aktivierungspotenzial aufwiesen, wenn sie mit einem Panel von GAL4-Treibern verwendet wurden. Um die In-vivo-Funktionalität zu demonstrieren, wurden diese Konstrukte dann verwendet, um zwei separate transgene An. gambiae-Linien durch PiggyBac-Transformation8 zu erzeugen: eine Treiberlinie mit GAL4, die von einem Mitteldarm-spezifischen Promotor angetrieben wird, und eine Responder-Linie, die sowohl die Luciferase- als auch die eYFP-Gene (Enhanced Yellow Fluorescent Protein) unter Regulation von UAS-Sequenzen enthält. Darmspezifische Luciferase-Aktivität und Fluoreszenz in den Nachkommen zeigten, dass das System in Anopheles effizient war. Seitdem wurden Treiberlinien erstellt, die Transgene in anderen Geweben exprimieren, die für die Vektorkapazität und insektizidresistenz wichtig sind, einschließlich Derozyten15 und Derhämozyten16, und in einem nahezu allgegenwärtigen Muster10. Zahlreiche UAS-Linien wurden auch erzeugt, um Gene zu untersuchen, von denen angenommen wird, dass sie an stoffwechsel- und sequestrierungsvermittelter Insektizidresistenz, kutikulärer Kohlenwasserstoffsynthese beteiligt sind, und um verschiedene Zell- und Gewebetypen fluoreszierend zu markieren (Tabelle 1). Für die Responder-Linien wird nun die ortsgerichtete Integration des Transgens durch ΦC31 katalysierten Rekombinationskassettenaustausch17,18 durchgeführt, um den genomischen Kontext der UAS-regulierten Gene zu fixieren. Auf diese Weise wird die Transgenexpression in Bezug auf die position der genomischen Insertion normalisiert, was einen genaueren Vergleich der phänotypischen Effekte verschiedener Kandidatengene ermöglicht.
Die bisher erstellten Responder-Linien sind so konzipiert, dass sie das Transgen entweder in erhöhten Konzentrationen exprimieren oder die Genexpression durch RNA-Interferenz (RNAi) reduzieren. Normalerweise werden cDNA-Klone mit der UAS-Sequenz fusioniert, um geeignete Expressionsplasmide zu erzeugen, aber auch vollständige genomische Sequenzen sind möglich, vorausgesetzt, sie sind nicht zu groß für das Klonen. Um Silencing-Konstrukte zu erzeugen, haben wir drei verschiedene Methoden verwendet, um geeignete tandeminvertierte Sequenzen zu erhalten, die Hairpin dsRNA bilden, die RNAi stimuliert. Dazu gehörten Fusions-PCR, asymmetrische PCR und kommerzielle Synthese von Haarnadelkonstrukten. Allen Methoden gemeinsam ist die Aufnahme einer Intronsequenz zwischen die invertierten Sequenzen, um die Klonstabilität zu gewährleisten. Es wurden Responder-Plasmide entwickelt, in die ein Gen von Interesse/RNAi-Konstrukt eingefügt werden kann15. Diese Plasmide tragen auch die erforderlichen ΦC31 attB-Stellen für RMCE (beschrieben in Adolfi begleitendem JoVE-Papier, das die RCME-Technik im Detail beschreibt). Protokolle, die die wichtigen Schritte abdecken, die bei der Auswahl der Sequenz für die Insertion in eines dieser Plasmide zur Überexpression erforderlich sind, sind in diesem Manuskript enthalten. Zusätzlich werden zwei Protokolle für die Erstellung von RNAi-Haarnadelkonstrukten beschrieben und veranschaulicht.
Bei der Schaffung neuer Linien ist die Identifizierung seltener transgener Individuen entscheidend für die Zucht, um transgene Kolonien zu etablieren und zu erhalten. Am wichtigsten für das GAL4-UAS-System ist, dass die Responder- und Treiberlinien unterschieden werden müssen, um Kreuze zu ermitteln und einzelne Nachkommen zu identifizieren, die beide Transgene tragen. Dies wird durch die Verwendung verschiedener dominanter selektorierbarer Markergene erreicht, die mit den Treiber- und Responder-Kassetten verknüpft sind. Am häufigsten handelt es sich dabei um fluoreszierende Markergene, die durch optische Filter (z.B. eYFP, eCFP, dsRed) deutlich unterscheidbar sind. Es ist wichtig, dass Marker in einem bekannten und zuverlässigen raumzeitlichen Muster ausgedrückt werden, da dies die Identifizierung von Anomalien und Kontaminationen erleichtert. Die Expression des fluoreszierenden Markergens wird routinemäßig durch den synthetischen 3xP3-Promotor reguliert, der in allen Stadien der Entwicklung von An. gambiae eine augen- und ventrale Ganglien-spezifische Expression verursacht19. Fluoreszierende Marker, die von 3xP3 kontrolliert werden, sind in allen in diesem Artikel beschriebenen Transformationsplasmiden enthalten. Ein Protokoll, das die gängigen Methoden zum Screening von fluoreszierenden An. gambiae Puppen GAL4-UAS-Linien beschreibt, ist hier enthalten.
Eines der Schlüsselelemente des GAL4-UAS-Systems ist die Notwendigkeit, die differentiell markierten Treiber- und Responder-Linien zu überqueren. Um dies zu tun, müssen Männchen und Weibchen aus jeder Linie vor der Paarung getrennt werden. Erwachsene sind durch das Sehen leicht unterscheidbar, aber für die Feststellung genetischer Kreuzungen ist es sinnvoll, die Geschlechter vor dem Auftauchen des Erwachsenen zu trennen, um sicherzustellen, dass keine Paarung stattgefunden hat. Der allgemeine Größenunterschied zwischen männlichen und weiblichen An. gambiae-Puppen ist zu variabel, um eine effiziente und zuverlässige Methode der Geschlechtsbestimmung zu sein20. Stattdessen liefern deutliche morphologische Unterschiede in den äußeren Genitalien eine verlässliche Grundlage für das Sexing bei An. gambiae. In diesem Artikel beschreiben wir eine zuverlässige Methode zum Sexing von An. gambiae Puppen, um geeignete Kreuzungen aufzustellen.
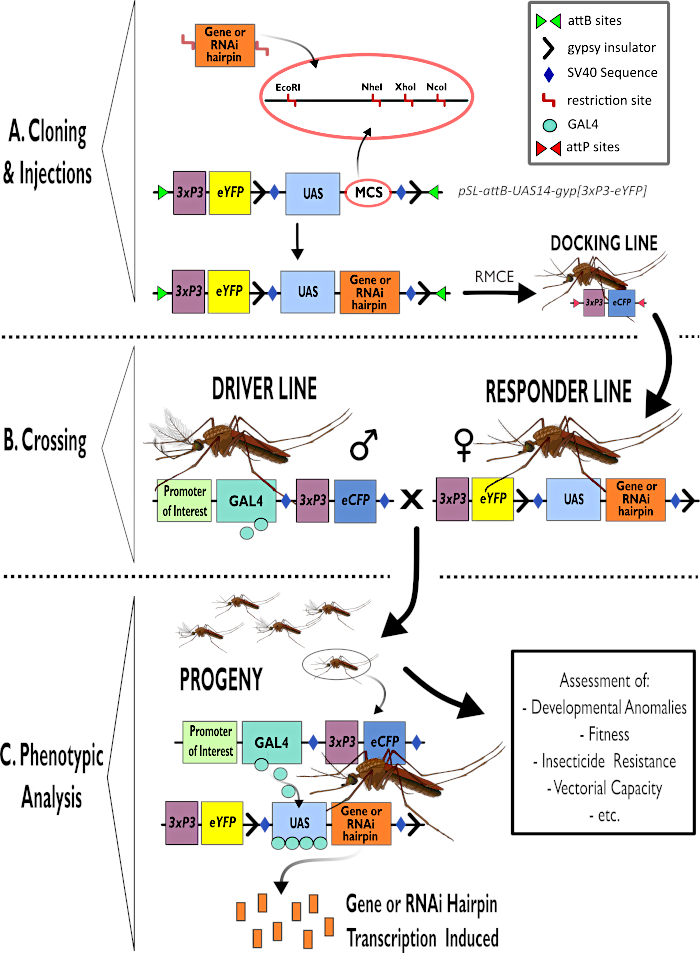
Abbildung 1 – Schematische Darstellung des Prozesses zur Verwendung des zweigliedrigen GAL4-UAS-Systems in Anopheles gambiae. (A) Die Hauptkomponenten eines Beispielvektors (pSL-attB-UAS14-gyp[3xp3-eYFP]) werden dargestellt, wobei die verfügbaren Restriktionsstellen (EcoRI, NheI, XhoI und NcoI) innerhalb der multiplen Klonierungsstellen aufgeführt sind, die für die Verwendung geeignet sind, das Haarnadelkonstrukt oder die kodierende Sequenz für das interessierende Gen einzufügen. Der Aufbau der Andocklinie ist ebenfalls dargestellt. (B) Der Kreuzungsschritt ist dargestellt und zeigt die Verwendung von Männchen aus der Fahrerlinie (mit GAL4-Fahrer durch einen Promoter von Interesse und eCFP, der vom 3xP3-Promoter gesteuert wird) und Weibchen aus der Responder-Linie (mit dem von einem UAS-Promotor kontrollierten Gen oder Haarnadelkonstrukt und einem eYFP-Marker, der vom 3xP3-Promotor kontrolliert wird). (C) Eine schematische Darstellung der GAL4-treibenden Expression des Gens, das für die Nachkommen des Kreuzes von Interesse ist, in B und eine Liste einiger der typischen Phänotypen, die bewertet werden. Abkürzungen: Multiple Cloning Site (MCS), Recombinase mediated cassette exchange (RMCE), Upstream Activator Sequence (UAS), enhanced yellow fluorescent protein (eYFP), enhanced cyan fluorescent protein (eCFP). Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Es ist die Verwendung von Kreuzen, die die zweigliedrige Natur des GAL4-UAS-Systems bietet, die gegenüber lineareren Ansätzen deutliche Vorteile bietet. Beispielsweise können viel mehr Kombinationen von Treiber- und Responder-Linien bewertet werden, als es möglich wäre, wenn für jede Promotor-Gen-Kombination eine neue transgene Linie erzeugt und aufrechterhalten werden müsste. Noch wichtiger ist, dass es die Analyse von Genen ermöglicht, die tödliche oder sterile Phänotypen produzieren, wenn ihre Expression gestört ist, die in einem linearen System schwer zu erzeugen / aufrechtzuerhalten sind. Solche tödlichen Phänotypen können sich in allen Entwicklungsstadien manifestieren, abhängig von der Genfunktion und der raumzeitlichen Expression, werden aber am häufigsten während der Embryonalentwicklung beobachtet. Die Visualisierung der Entwicklung von Mückenembryonen erfordert die Reinigung des undurchsichtigen Chorions, das die Eier bedeckt. Nach Methoden, die in Trpiš (1970) 21 und Kaiser et al. (2014) 22 beschrieben sind, beschreiben wir die Protokolle, die wir verwenden, um Embryonen unter Beibehaltung der strukturellen Integrität zu fixieren, und Bleichen, um die Endochorion zu beseitigen, die eine mikroskopische Visualisierung und Bildgebung ermöglicht.
