In 1998, Fire en Mello meldde dat dubbelstrengs RNA (dsRNA) remming van genfunctie kan induceren in Caenorhabditis elegans 1. Deze reactie veroorzaakt door dsRNA werd genoemd RNA interferentie (RNAi) en dergelijke RNAi-gemedieerde silencing werd gerapporteerd als zijnde geconserveerd in dieren, planten en schimmels 2-7. In planten en sommige dieren, RNAi functies systemisch, wat betekent dat het effect kan zich verspreiden naar andere cellen / weefsels waarbij dsRNA niet rechtstreeks geïntroduceerd (beoordeeld in 8-10). Wetenschappers hebben gebruik gemaakt van deze endogene cellulaire RNAi respons door het ontwerpen van dsRNA's om genen van belang gericht zijn gemaakt, waardoor de functie van genen neerhalen zonder direct het manipuleren van het genoom (beoordeeld in 11-14).
RNAi is een krachtig hulpmiddel voor functionele studies vanwege de volgende voordelen. Eerst, zelfs met minimale informatie gensequentie, een gen kan worden gevist met RNAi. Dit is vooral belangrijk voor studies van niet-model organismen ontbreekt genomische of transcriptomics data. Ten tweede, in organismen waar RNAi respons robuust systemische RNAi-gemedieerde gen knockdown kan worden uitgevoerd op bijna elk ontwikkelingsstadium. Deze functie is zeer nuttig voor het bestuderen van de functie van pleiotrope genen. Ten derde, in sommige gevallen, RNAi effecten uitbreiden tot de gonaden en nageslacht, zodat fenotypes worden waargenomen bij nakomelingen 15,16. Dit fenomeen, bekend als ouderlijke RNAi (pRNAi), is bijzonder voordelig voor genen beïnvloeden embryonale ontwikkeling, talrijke nakomelingen van één ouder geïnjecteerd zonder directe manipulatie eieren kunnen worden onderzocht. Daarom pRNAi is de voorkeurswerkwijze. Indien pRNAi ineffectief, bijvoorbeeld voor genen die nodig zijn voor oogenesis, dan embryonale RNAi (eRNAi) worden gebruikt. Ten vierde kan RNAi worden gebruikt om het equivalent van een allelische reeks, dat de hoeveelheid dsRNA geleverd over een bereik kan worden gevarieerd wekken zwakke produceren sterke gebreken. Dergelijke gradatie van fenotypes kan nuttig zijn voor het begrijpen van genfunctie wanneer het gen is betrokken bij een complex proces en / of volledig verlies van functie dodelijk. Ten vijfde, de levering van dsRNA is over het algemeen gemakkelijk en haalbaar is, met name in dieren die robuuste systemische RNAi reacties. dsRNA kan worden ingevoerd door micro-injectie 1,5, het voeden / inname 17,18, weken, 19,20 en virus / bacteriën gemedieerde aflevering 21,22. Zesde, in tegenstelling tot sommige gentargeting / bewerken werkwijzen hoeft te screenen op organismen die de mutatie of genetische kruisingen aan homozygoten genereren bij gebruik RNAi uitvoeren. Daarom in vergelijking met vele andere technieken voor het bestuderen van genfunctie, RNAi is snel, goedkoop en kunnen worden toegepast voor grootschalige schermen 23-25.
De brede toepasbaarheid van RNAi verschaft middelen functionele studies uit te voeren in een breed scala van organismen, het uitbreiden van het bereik van de soorten beschikbaar voor studie beyond de traditionele modelsystemen waarvoor genetische instrumenten ontwikkeld. Bijvoorbeeld, studies met niet-modelsystemen vereist om inzicht in de evolutie van genen en gen netwerken geven door vergelijking van de functies van orthologen voor soorten van verschillende modes of ontwikkeling vertonen duidelijke morfologische kenmerken 26-29. Deze types van studies zullen een beter begrip van de biologische diversiteit te voorzien, met gevolgen voor zowel toegepast en fundamenteel onderzoek.
Omdat het grootste dier groep op de planeet, insecten zorgen voor een geweldige kans om de mechanismen te onderzoeken onderliggende diversiteit. Bovendien, insecten zijn over het algemeen klein, hebben korte levenscycli, hoge vruchtbaarheid, en zijn eenvoudig aan de achterzijde in het lab. In de afgelopen twee decennia, is RNAi is met succes toegepast bij insecten verspreid over orders, met inbegrip van Diptera (true vliegen) 5, Lepidoptera (vlinders en motten) 30, Coleoptera (kevers) 16,31, Hymenoptera (sawfleugens, wespen, mieren en bijen) 32, Hemiptera (wantsen), Isoptera (termieten) 34, Blattodea (kakkerlakken) 35, Orthoptera (krekels, sprinkhanen, sprinkhanen, en katydids) 36 en Phthiraptera (luizen) 37. Succesvolle toepassing van RNAi heeft verstrekt functionele gegevens voor onderzoek naar patronen in de vroege embryogenese (anterior-posterior as 32, dorsale-ventrale as 28, segmentatie 26,38), geslachtsbepaling 39,40, chitine / cuticula biosynthese 41, ecdyson signalering 42, sociaal gedrag 43, en nog veel meer. RNAi methoden ontwikkeld voor verschillende insectensoorten kunnen bijkomende voordeel dat zij mogelijk bruikbaar voor ongediertebestrijding (herzien 44-46) te zijn. RNAi effecten genspecifieke en soortspecifiek zijn, zolang als niet-geconserveerde gebieden worden gekozen voor targeting. Voor nuttige insecten zoals bijen en zijderupsen, gericht op genen die van vitaal belang voor het voortbestaan vanvirussen of parasieten om besmetting te bestrijden kan een nieuwe strategie om deze soorten te beschermen 47,48 bieden.
Dermestes Maculatus (D. maculatus), gangbare naam hide kever, wordt wereldwijd verspreid, met uitzondering van Antarctica. Als holometabolous insect, de D. maculatus levenscyclus omvat embryonale, larven, pupal en volwassen stadia (figuur 1). Omdat het voedt zich met vlees, wordt D. maculatus gebruikt in musea dode dieren schets maken en forensische entomologen kunt het gebruiken om het tijdstip van overlijden 49,50 te schatten. D. maculatus zich voedt met dierlijke producten zoals kadavers, gedroogd vlees, kaas, en de poppen / cocons van andere insecten en veroorzaakt daarmee schade aan huishoudens, opgeslagen voedsel, en de zijde, kaas en vlees-industrie 51,52. Het toepassen van RNAi in deze kever zou een efficiënte en milieuvriendelijke manier om de economische gevolgen te minimaliseren. Ons laboratorium heeft gebruikt D. maculatus als een nieuwe model insect segmentering 53 bestuderen. Naast het feit dat vatbaar lab fokken, D. maculatus van belang voor fundamenteel onderzoek omdat het een tussenproduct kiem ontwikkelaar, waardoor het een nuttige soorten naar de overgang tussen korte en lange kiem ontwikkeling bestuderen.
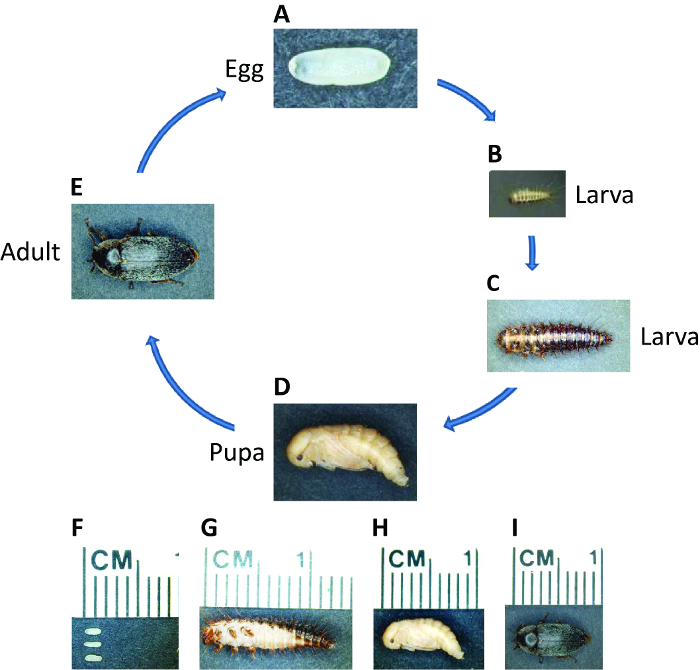
Figuur 1: Life Cycle van D. maculatus. Foto's van D. maculatus in verschillende levensfasen, zoals aangegeven. De levenscyclus van ei tot adult bedraagt drie weken bij 30 ° C, maar meer bij lagere temperaturen. (A, F) Vers gelegd embryo's zijn wit tot lichtgeel en ovaal, ongeveer 1,5 mm in lengte. Embryogenese draait ~ 55 uur bij 30 ° C. (B, C en G) Larven hebben donkere gepigmenteerde strepen en zijn bedekt met borstels. Larven gaan door verschillende stadia, afhankelijk van de omgeving en hun lengte kan zich uitstrekken tot meer dan 1 cm. (D, H) </strong> Young poppen zijn lichtgeel. Verpopping vindt ~ 5-7 dagen bij 30 ° C. (E, I) Kort na eclosion, verschijnt donkere pigmentvlekken op de volwassen kever lichaam. Volwassenen kunnen leven tot enkele maanden en een vrouwtje kan honderden embryo's lag over haar leven. Klik hier om een grotere versie van deze figuur te bekijken.
Eerder toonden we aan dat RNAi is effectief in het neerhalen gen-functie in D. maculatus 53. Hier onze ervaring fokken D. maculatus kolonies in het laboratorium wordt gedeeld met stap-voor-stap protocollen voor zowel embryonale en ouderlijke RNAi set-up, injectie, na de injectie zorg en fenotypische analyse. De dsRNA-gemedieerde gen knockdown en analysemethoden hier geïntroduceerd bieden niet alleen gedetailleerde informatie voor de aanpak van vraagstukken in D. maculatus, maar ook potentiële betekenis for toepassing van RNAi in andere niet-model kever / insectensoorten.
