Nel 1998, Fuoco e Mello hanno riferito che a doppio filamento di RNA (dsRNA) può indurre inibizione della funzione del gene in Caenorhabditis elegans 1. Questa risposta innescato da dsRNA è stato chiamato interferenza dell'RNA (RNAi), e tale silenziamento genico RNAi-mediata è stato segnalato per essere conservato in animali, piante e funghi 2-7. In piante e alcuni animali, funzioni RNAi sistemica, il che significa che l'effetto può diffondersi ad altre cellule / tessuti dove dsRNA non viene introdotto direttamente (rivisto nel 8-10). Gli scienziati hanno fatto uso di questo endogena risposta RNAi cellulare progettando dsRNAs di indirizzare i geni di interesse, battendo così giù la funzione del gene senza manipolare direttamente il genoma (rivisto nel 11-14).
RNAi è un potente strumento per studi funzionali causa dei seguenti vantaggi. In primo luogo, anche con il minimo informazioni sequenza del gene, un gene può essere mirata utilizzando RNAi. Ciò è particolarmente importante per studies di organismi non-modello privi di dati genomici e trascrittomica. In secondo luogo, negli organismi dove la risposta RNAi è robusta sistemica RNAi mediata knockdown gene può essere effettuata in quasi ogni stadio di sviluppo. Questa funzione è molto utile per studiare la funzione dei geni pleiotropici. In terzo luogo, in alcuni casi, effetti RNAi diffuso ai gonadi e discendenti, in modo tale che i fenotipi si osservano nella prole 15,16. Questo fenomeno, noto come genitori RNAi (pRNAi), è particolarmente vantaggioso per i geni che influenzano lo sviluppo embrionale, come numerosa prole prodotta da un solo genitore iniettato può essere esaminato, senza manipolazione diretta di uova. Per queste ragioni, pRNAi è il metodo di scelta. Tuttavia, se pRNAi è inefficace, ad esempio per i geni necessari per la oogenesi, allora embrionale RNAi (eRNAi) deve essere utilizzato. Quarto, RNAi può essere utilizzato per generare l'equivalente di una serie allelica dal fatto che la quantità di dsRNA consegnato può essere variata su un intervallo di produrre debole per difetti forti. Tale gradazione di fenotipi può essere utile per comprendere la funzione del gene quando il gene è coinvolto in un processo complesso e / o completa perdita di funzione è letale. In quinto luogo, la consegna di dsRNA è generalmente facile e fattibile, soprattutto negli animali che mostrano solide risposte RNAi sistemiche. dsRNA può essere introdotto da microiniezione 1,5, alimentazione / ingestione 17,18, ammollo, 19,20 e il virus / batteri-mediata consegna 21,22. Sesto, a differenza di alcuni metodi di targeting / modifica dei geni, non vi è alcuna necessità di screening per gli organismi che trasportano la mutazione o di effettuare incroci genetici per generare omozigoti quando si usa RNAi. Pertanto, rispetto a molte altre tecniche per studiare la funzione del gene, RNAi è veloce, poco costoso, e può essere applicato per schermi di grandi dimensioni 23-25.
L'ampio programma di utilità di RNAi fornisce i mezzi per effettuare studi funzionali in una vasta gamma di organismi, ampliando la gamma di specie disponibili per lo studio beyond i sistemi modello tradizionali per i quali sono stati sviluppati strumenti genetici. Ad esempio, gli studi che utilizzano sistemi non-modello sono tenuti a dare intuizioni l'evoluzione dei geni e reti geniche confrontando le funzioni di ortologhi da specie che rappresentano diverse modalità di sviluppo o esporre distinte caratteristiche morfologiche 26-29. Questi tipi di studi forniranno una migliore comprensione della diversità biologica, con conseguenze per la ricerca sia applicata e di base.
Essendo il più grande gruppo di animali sul pianeta, gli insetti offrono una grande opportunità per esplorare i meccanismi alla base della diversità. Inoltre, gli insetti sono in genere piccole, hanno cicli di vita brevi, ad alta fecondità, e sono facili da dietro in laboratorio. Negli ultimi due decenni, RNAi è stato applicato con successo negli insetti che abbracciano gli ordini, tra cui Ditteri (vere mosche) 5, lepidotteri (farfalle e falene) 30, Coleotteri (coleotteri) 16,31, Imenotteri (sawfbugie, vespe, formiche e api) 32, Hemiptera (veri insetti), Isotteri (termiti) 34, Blattodea (scarafaggi) 35, ortotteri (grilli, cavallette, locuste e grilli) 36 e Phthiraptera (pidocchi) 37. Applicazione di successo di RNAi ha fornito dati funzionali per gli studi di patterning in embriogenesi precoce (asse anteriore-posteriore 32, dorso-ventrale asse 28, la segmentazione 26,38), determinazione del sesso, 39,40 chitina / cuticola biosintesi 41, ecdysone segnalazione 42, comportamento sociale 43, e altro ancora. Metodi RNAi sviluppati per diverse specie di insetti possono essere di ulteriore vantaggio in quanto sono suscettibili di essere utile per il controllo dei parassiti (rivisto nel 44-46). effetti RNAi saranno nonché specie-specifico gene-specifico, purché regioni non conservate sono scelti per il targeting. Per le specie di insetti utili come le api e bachi da seta, di targeting dei geni vitali per la sopravvivenza divirus o parassiti per controllare l'infezione possono fornire una nuova strategia per proteggere queste specie 47,48.
Dermestes maculatus (D. maculatus), comune nome nascondere coleottero, è distribuito in tutto il mondo tranne che per l'Antartide. Come un insetto olometaboli, il ciclo di vita D. maculatus comprende embrionale, larvale, pupa, e stadi adulti (Figura 1). Perché si nutre di carne, D. maculatus è utilizzato nei musei per skeletonize animali morti e entomologi forensi può essere utilizzato per stimare momento della morte 49,50. D. maculatus si nutre di prodotti di origine animale, tra cui carcasse, carne secca, formaggio e le pupe / bozzoli di altri insetti e provoca così danni alle famiglie, alimenti conservati, e la seta, il formaggio, e industrie della carne 51,52. L'applicazione di RNAi in questo coleottero potrebbe fornire un modo efficiente ed ecologico per ridurre al minimo il suo impatto economico. Il nostro laboratorio ha utilizzato D. maculatus come nuovo model insetto per studiare la segmentazione 53. Oltre ad essere suscettibili di laboratorio allevamento, D. maculatus è di interesse per la ricerca di base in quanto è uno sviluppatore intermedio di germi, che lo rende una specie utili per studiare la transizione tra breve e di sviluppo a lungo germe.
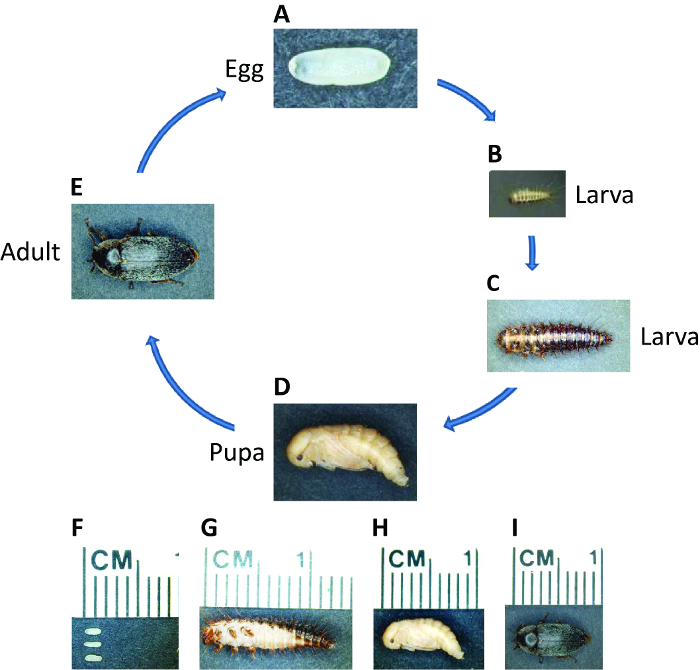
Figura 1: Ciclo di vita di D. maculatus. Fotografie di D. maculatus in diverse fasi della vita, come indicato. Il ciclo di vita da uovo ad adulto dura tre settimane a 30 ° C, ma più a lungo a temperature più basse. (A, F) embrioni appena deposte sono di colore bianco a giallo chiaro e ovali, circa 1,5 mm di lunghezza. Embriogenesi prende ~ 55 ore a 30 ° C. (B, C e G) larve hanno le strisce pigmentate scure e sono ricoperte di setole. Le larve passano attraverso diversi stadi a seconda dell'ambiente e la loro lunghezza può estendersi fino a oltre 1 centimetro. (D, H) </strong> Giovani pupe sono giallo chiaro. Pupation prende ~ 5 – 7 giorni a 30 ° C. (E, I) Poco dopo eclosion, pigmentazione scura appare sopra il corpo adulto coleottero. Gli adulti possono vivere fino a diversi mesi e una femmina può deporre centinaia di embrioni sopra il suo ciclo di vita. Clicca qui per vedere una versione più grande di questa figura.
In precedenza, abbiamo dimostrato che RNAi è efficace nel abbattendo la funzione del gene in D. maculatus 53. Qui la nostra esperienza di allevamento colonie D. maculatus in laboratorio è condiviso con i protocolli passo-passo sia embrionale e genitori RNAi set-up, iniezione, cura post-iniezione, e l'analisi fenotipica. Il silenziamento genico e analisi metodi dsRNA-mediate introdotti qui non solo di fornire informazioni dettagliate per affrontare questioni in D. maculatus, ma hanno anche un significato potenziale foR applicazione di RNAi in altre specie non-modello scarabeo / insetto.
