En 1998, le feu et Mello ont rapporté que l' ARN double brin (ARNdb) peut induire l' inhibition de la fonction des gènes dans Caenorhabditis elegans 1. Cette réponse déclenchée par l' ARN double brin a été appelé ARN interférence (ARNi), et un tel silençage génique à médiation par ARNi a été rapportée être conservée chez les animaux, les plantes et les champignons de 2-7. Chez les plantes et certains animaux, les fonctions ARNi systémique, ce qui signifie que l'effet peut se propager à d' autres cellules / tissus où ARNdb est pas directement introduit (revue dans 8-10). Les scientifiques ont fait usage de cette réponse ARNi cellulaire endogène en concevant ARNdb pour cibler des gènes d'intérêt, ce qui frappe vers le bas la fonction du gène sans manipuler directement le génome (revue dans 11-14).
L'ARNi est un outil puissant pour des études fonctionnelles en raison des avantages suivants. Tout d'abord, même avec des informations sur la séquence du gène minimum, un gène peut être ciblé en utilisant l'ARNi. Ceci est particulièrement important pour les rudies d'organismes non-modèles dépourvus de données génomiques ou transcriptomiques. D'autre part, dans les organismes où la réponse ARNi est robuste systémique, knock-down de gènes à médiation par ARNi peut être réalisée à pratiquement n'importe quel stade de développement. Cette fonctionnalité est très utile pour étudier la fonction des gènes pléiotropiques. Troisièmement, dans certains cas, les effets ARNi se propagent aux gonades et descendants, tels que les phénotypes sont observés chez la progéniture 15,16. Ce phénomène, connu sous le nom ARNi parental (pRNAi), est particulièrement avantageux pour les gènes impact sur le développement embryonnaire, aussi nombreux descendants produits par un seul parent injecté peut être examinée sans manipulation directe des œufs. Pour ces raisons, pRNAi est la méthode de choix. Toutefois, si pRNAi est inefficace, par exemple pour les gènes nécessaires à l'ovogenèse, puis embryonnaires ARNi (eRNAi) doit être utilisé. Quatrièmement, l'ARNi peut être utilisé pour générer l'équivalent d'une série allélique en ce que la quantité d'ARNdb livré peut varier sur une plage pour produire faible pour les défauts forts. Une telle gradation des phénotypes peut être utile pour la compréhension de la fonction des gènes lorsque le gène est impliqué dans un processus complexe et / ou la perte complète de la fonction est létale. Cinquièmement, la livraison d'ARNdb est généralement facile et réalisable, en particulier chez les animaux montrant robustes réponses systémiques ARNi. ARNdb peut être introduit par microinjection 1,5, l' alimentation / l' ingestion 17,18, 19,20 trempage et le virus de la livraison / bactéries médiée 21,22. Sixième, à la différence des méthodes de ciblage / d'édition de gènes, il n'y a pas besoin de dépister les organismes porteurs de la mutation ou de procéder à des croisements génétiques pour générer des homozygotes lors de l'utilisation d'ARNi. Par conséquent, par rapport à de nombreuses autres techniques pour étudier la fonction des gènes, l' ARNi est rapide, peu coûteuse et peut être appliqué pour les écrans de grande envergure 23-25.
La grande utilité de l'ARNi fournit des moyens de réaliser des études fonctionnelles dans un large éventail d'organismes, en élargissant la gamme des espèces disponibles pour l'étude beyond les systèmes modèles traditionnels pour lesquels des outils génétiques ont été développés. Par exemple, des études utilisant des systèmes non-modèle sont tenus de donner un aperçu de l'évolution des gènes et des réseaux de gènes en comparant les fonctions de orthologues d'espèces représentant différents modes de développement ou présentant des caractéristiques morphologiques distinctes 26-29. Ces types d'études fourniront une meilleure compréhension de la diversité biologique, avec des impacts de la recherche à la fois fondamentale et appliquée.
Étant le plus grand groupe d'animaux sur la planète, les insectes fournissent une excellente occasion d'explorer les mécanismes de la diversité sous-jacente. En outre, les insectes sont généralement de petite taille, ont des cycles de vie courts, une fécondité élevée, et sont faciles à élever en laboratoire. Au cours des deux dernières décennies, l' ARNi a été appliquée avec succès dans les insectes couvrant les commandes, y compris les diptères (mouches vraies) 5, lépidoptères (papillons et mites) 30, coléoptères (scarabées) 16,31, hyménoptères (Sawfmensonges, les guêpes, les fourmis et les abeilles) 32, Hemiptera (punaises), isoptères (termites) 34, Blattodea (cafards) 35, orthoptères (grillons, sauterelles, criquets et sauterelles) 36 et Phthiraptera (poux) 37. L' application réussie de l' ARNi a fourni des données fonctionnelles pour les études de structuration dans l' embryogenèse précoce (antéro-postérieur axe 32, dorso-ventral axe 28, segmentation 26,38), la détermination du sexe 39,40, chitine / cuticule biosynthèse 41, ecdysone de signalisation 42, comportement social 43, et plus encore. Méthodes d' ARNi développées pour différentes espèces d'insectes peuvent être d'avantage supplémentaire en ce sens qu'elles sont susceptibles d'être utiles pour la lutte antiparasitaire (passé en revue dans 44-46). les effets d'ARNi seront aussi bien que spécifique de l'espèce spécifique du gène, aussi longtemps que les régions non conservées sont choisies pour le ciblage. Pour les espèces d'insectes bénéfiques comme les abeilles et les vers à soie, en ciblant les gènes vitaux pour la survie devirus ou des parasites pour contrôler l' infection peuvent fournir une nouvelle stratégie visant à protéger ces espèces 47,48.
Dermestes maculatus (D. maculatus), nom commun hide scarabée, est distribué dans le monde entier à l' exception de l' Antarctique. Comme un insecte holométabole, le cycle de vie D. maculatus comprend embryonnaire, larvaire, pupe et stades adultes (Figure 1). Parce qu'il se nourrit de chair, D. maculatus est utilisé dans les musées à squelettage animaux morts et entomologistes médico – légale peut l' utiliser pour estimer le temps de la mort 49,50. D. maculatus se nourrit de produits d'origine animale , y compris les carcasses, la viande séchée, du fromage et des pupes / cocons d'autres insectes et provoque ainsi des dommages aux ménages, les aliments stockés, et la soie, le fromage et les industries de la viande 51,52. L'application de l'ARNi dans ce coléoptère pourrait fournir un moyen efficace et respectueux de l'environnement afin de minimiser son impact économique. Notre laboratoire a utilisé D. maculatus comme une nouvelle model insectes pour étudier la segmentation 53. En plus d'être prête à l' élevage de laboratoire, D. maculatus est d'intérêt pour la recherche fondamentale car elle est un développeur intermédiaire germe, ce qui en fait une espèce utile pour étudier la transition entre court et le développement à long germe.
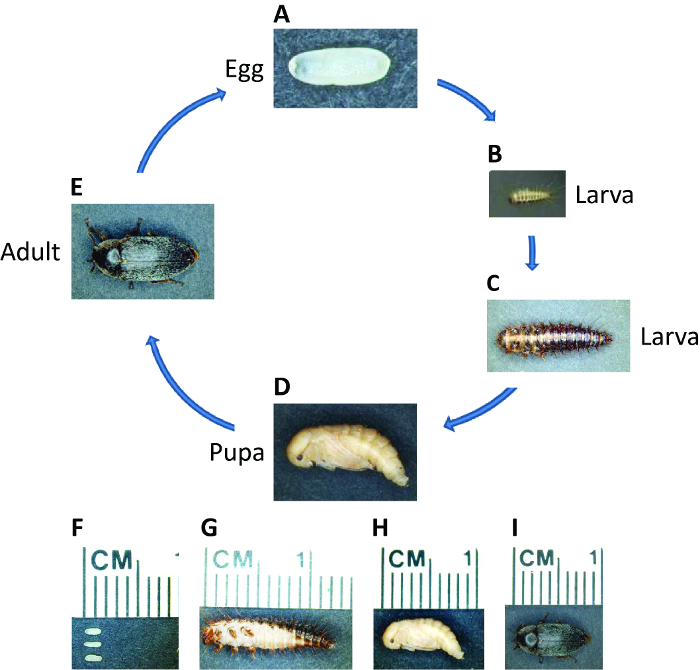
Figure 1: Cycle de vie de D. maculatus. Photographies de D. maculatus à différents stades de la vie, comme indiqué. Le cycle de vie de l'œuf à l'adulte dure trois semaines à 30 ° C mais plus à des températures plus basses. (A, F) des embryons fraîchement pondus sont blancs à jaune clair et ovales, d' environ 1,5 mm de longueur. Embryogenèse prend ~ 55 heures à 30 ° C. (B, C et G) Larves ont des rayures pigmentées sombres et sont recouverts de soies. Les larves passent par plusieurs stades en fonction de l'environnement et leur longueur peut aller jusqu'à plus de 1 cm. (D, H) </strong> Jeune pupes sont jaune clair. La nymphose a ~ 5 – 7 jours à 30 ° C. (E, I) Peu de temps après l' éclosion, la pigmentation foncée apparaît sur le corps de coléoptère adulte. Les adultes peuvent vivre jusqu'à plusieurs mois et une femelle peut pondre des centaines d'embryons au cours de sa vie. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
Auparavant, nous avons montré que l' ARNi est efficace pour abattre la fonction du gène dans D. maculatus 53. Voici notre expérience d' élevage colonies D. maculatus dans le laboratoire est partagé avec les protocoles étape par étape pour RNAi set-up à la fois embryonnaire et parental, l' injection, les soins post-injection, et l' analyse phénotypique. Les knockdown et méthodes d' analyse de gènes ARNdb médiées introduits ici non seulement fournir des informations détaillées pour aborder les questions de D. maculatus, mais aussi avoir une signification potentielle for l'application de l'ARNi dans d'autres espèces d'insectes / non-modèle coléoptères.
