1998年には、火とメロは、二本鎖RNA(dsRNA)が線虫(Caenorhabditis elegans)1における遺伝子機能の抑制を誘導することができることを報告しました。該dsRNAによって誘発この応答は、RNA干渉(RNAi)と命名し、そしてそのようなRNAiを介した遺伝子サイレンシングは、動物、植物、および真菌2-7に保存されることが報告されました。植物や一部の動物において、RNAi機能は、全身、意味の効果が(8月10日に概説)のdsRNAが直接導入されていない他の細胞/組織に広がることができます。科学者たちは、それによって直接(11月14日に見直さ)ゲノムを操作することなく、遺伝子機能をノックダウン、目的の遺伝子を標的とするためのdsRNAを設計することにより、この内因性細胞RNAi応答を利用してきました。
RNAiは、次のような利点のために機能的研究のための強力なツールです。まず、偶数最小限の遺伝子配列情報を用いて、遺伝子は、RNAiを使用して標的化することができます。これは、STのために特に重要ですゲノムやトランスクリプトームデータを欠く非モデル生物のudies。第二に、RNAi応答がロバスト全身である生物で、RNAiを介した遺伝子ノックダウンは、ほぼすべての発達段階で行うことができます。この機能は、多面的な遺伝子の機能を研究するために非常に有用です。第三に、いくつかの場合には、RNAi効果を表現型子孫15,16において観察されるように、生殖腺および子孫に広がります。単一注入親によって生成多数の子孫は、卵を直接操作することなく検査することができるように、親のRNAi(pRNAi)として知られるこの現象は、胚発生に影響を与える遺伝子について特に有利です。これらの理由から、pRNAiを選択する方法です。 pRNAiが無効である場合は、卵形成のために必要な遺伝子のために、たとえば、その後、胚のRNAi(eRNAi)を使用する必要があります。 dsRNAの量は強い欠陥に弱い生成する範囲にわたって変化させることができる送達という点で第四に、RNAiは、対立遺伝子シリーズの等価物を生成するために使用することができます。表現型のようなグラデーションは、遺伝子は致命的である複雑なプロセスおよび/または機能の完全な喪失に関与しているときに遺伝子機能を理解するのに役立つことができます。第五に、dsRNAの配信は、特に堅牢な全身のRNAi応答を示す動物では、一般的に簡単で、実現可能です。 dsRNAは、マイクロインジェクション1,5、摂食/摂取17,18、浸漬、19,20およびウイルス/細菌媒介送達21,22によって導入することができます。第六に、いくつかの遺伝子ターゲティング/編集方法とは異なり、変異を有する生物をスクリーニングするまたはRNAiを用いた場合にホモ接合体を生成するための遺伝的交雑を行う必要はありません。したがって、遺伝子機能を研究するための多くの他の技術と比較して、RNAiは、速く、安価であり、かつ大規模なスクリーニング23-25に適用することができます。
RNAiの広範なユーティリティがbeyon研究のために利用可能な種の範囲を拡大し、生物の広い範囲で機能的研究を遂行するための手段を提供します遺伝的ツールが開発されているために、従来のモデルシステムをdは。例えば、非モデル系を用いた研究は、異なる開発モードを表すか、別個の形態学的特徴26〜29を示す種由来のオルソログの機能を比較することにより、遺伝子および遺伝子ネットワークの進化への洞察を得るために必要とされます。研究のこれらのタイプは、印加されたと基本の両方の研究のための影響で、生物多様性のより良い理解を提供します。
地球上で最大の動物群である、昆虫は多様性のメカニズムを探求する絶好の機会を提供します。また、昆虫は、一般的に小さく、短いライフサイクル、高い繁殖力を持っている、と研究室の後部に簡単です。過去20年間において、RNAiが正常に双翅目(真のハエ)5を含む、注文にまたがる昆虫で適用されてきた、鱗翅目(蝶や蛾)30、鞘翅目(甲虫)16,31、膜翅目(sawf嘘、ハチ、アリやハチ)32、半翅目(真のバグ)、等翅目(シロアリ)34、Blattodea(ゴキブリ)35、直翅目(コオロギ、バッタ、イナゴ、とキリギリス)36と、シラミ目(シラミ)37。 RNAiの成功のアプリケーションは、エクジソンが42シグナル伝達、初期胚発生におけるパターン形成の研究のための機能データ(前後軸32、背腹軸28、セグメンテーション26,38)、性別決意39,40、キチン/キューティクルの生合成41を提供してきました社会的行動43、およびより多くの。彼らは(44-46に概説)害虫防除のために有用である可能性があるという点で異なる昆虫種のために開発されたRNAi法は、付加的な有益であり得ます。 RNAi効果は、非保存領域を標的とするために選択されている限り、遺伝子特異的なだけでなく、種特異的であろう。の生存に不可欠な遺伝子を標的ミツバチやカイコなどの有益な昆虫種については、感染を制御するためのウイルスまたは寄生虫は、これらの種47,48を保護するための新規な戦略を提供することができます。
Dermestes maculatus(D. maculatus)、一般名の非表示カブトムシは、南極大陸を除く世界的に分布しています。完全変態の昆虫としては、D. maculatusのライフサイクルは、胚、幼虫、さなぎ、および成体期( 図1)が含まれています。それは肉を餌にするので、D. maculatusは死んだ動物を骸骨にするために美術館に使用され、法医学昆虫学者は、死亡49,50の時間を推定するためにそれを使用することができます。 D. maculatusは死体、乾燥肉、チーズ、および他の昆虫の蛹/繭などの動物性食品に供給し、このように家計へのダメージ、保存された食品、シルク、チーズ、肉産業51,52の原因となります。この甲虫でRNAiを適用すると、その経済的影響を最小化するための効率的かつ環境に優しい方法を提供することができます。私たちの研究室では、新しいメートルとしてD. maculatusを使用していますセグメンテーション53を研究するodel昆虫。それはそれの有用な種は短期および長期の生殖発達の間の移行を検討すること、中間胚の開発者であるとして、実験室飼育の影響を受けやすいことに加えて、D. maculatusは、基礎研究のために重要です。
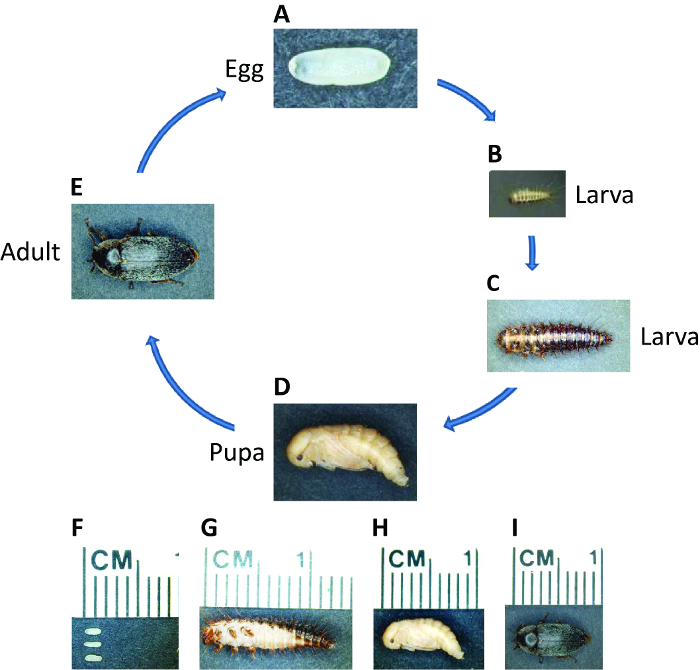
図1:D. maculatusのライフサイクル。示されているように異なるライフステージでのD. maculatusの写真、。卵から大人までのライフサイクルは、より低い温度でより長い30℃で3週間かかりますが。 (A、F)産まれたての胚は、長さ約1.5ミリメートル、淡黄色の楕円形に白です。胚は〜30℃で55時間かかります。 (B、C、およびG)幼虫が暗く着色されたストライプを有しており、毛で覆われています。幼虫は、環境に応じて、いくつかの齢を通過し、その長さは1オーバーセンチにまで拡張することができます。 (D、H) </strオング>ヤング蛹は淡黄色です。 30℃で7日間 – 蛹化は〜5かかります。 (E、I)間もなく羽化後、暗い色素沈着は、大人のカブトムシの体の上に表示されます。大人数ヶ月まで生きることができ、一人の女性が生涯にわたって胚の数百人を置くことができます。 この図の拡大版をご覧になるにはこちらをクリックしてください。
以前、我々は、RNAiは、D. maculatus 53における遺伝子機能のノックダウンに有効であることが示されました。ここでは実験室でD. maculatusコロニーを飼育経験は、胚と親の両方のRNAiセットアップ、注射、注射後のケア、および表現型解析のためのステップバイステップのプロトコルと一緒に共有されています。 D. maculatusで質問に対処するための詳細な情報を提供するだけでなく、foの潜在的な重要性を持っているだけでなく、ここで紹介したdsRNA媒介遺伝子ノックダウンと分析方法rの他の非モデル甲虫/昆虫種でRNAiを適用します。
