Pseudomonas aeruginosa (Pa) является оппортунистическим патогеном, который устанавливает хронические инфекции у людей с ослабленным иммунитетом. Для людей с генетическим заболеванием муковисцидоз (МВ) эти инфекции могут охватывать всю жизнь. Муковисцидоз вызывает накопление вязкой, богатой питательными веществами мокроты в дыхательных путях, которая со временем колонизируется различными микробными патогенами. Па является одним из наиболее распространенных патогенов муковисцидоза, колонизируя дыхательные пути в раннем детстве и устанавливая трудноизуемые инфекции1. Па остается значительной клинической проблемой и считается ведущей причиной смертности у лиц с муковисцидозом, несмотря на улучшение схем терапии в последниегоды2,3. Этот фенотип персистенции и повышение толерантности к антибиотикам принесли Па место в группе патогенов, идентифицированных как Центрами по контролю и профилактике заболеваний (CDC), так и Всемирной организацией здравоохранения (ВОЗ) в качестве исследовательских приоритетов для разработки новых терапевтических стратегий – патогенов ESKAPE4.
Как и другие патогены ESKAPE, приобретенная устойчивость к антибиотикам распространена в Pa,но есть также много внутренних свойств, которые способствуют антимикробной толерантности Pa. Среди них способность Па образовывать агрегаты — высокоплотные кластеры из ~10-1000 клеток, которые могут наблюдаться при множественных инфекциях, в том числе при МВ мокроте больного5,6. Подобно Pa, изученным в других системах биопленки, агрегаты Pa демонстрируют клинически значимые фенотипы, такие как повышенная устойчивость к антибиотикам и активация клеточно-клеточной коммуникации (кворум-зондирование (QS)). Например, было показано, что агрегаты Па используют QS-регулируемое поведение для борьбы с другими микробами, а также переносят антимикробные методы лечения, такие как производство пиоцианина7. Способность изучать такое поведение дает захватывающее представление о бактериальных экосистемах в среде, подобной той, в которой они существуют в организме человека.
Одной из самых больших проблем при изучении того, как агрегаты Pa реагируют на изменение среды мокроты, является отсутствие питательных и надежных систем, которые способствуют образованию агрегатов. Многое из того, что известно о Па, было обнаружено с использованием систем in vitro, в которых клетки растут планктонально или в характерной поверхностно-прикрепленной, «грибной» архитектуре, которая не наблюдалась in vivo8. В то время как классические модели роста биопленки, такие как прототочные клетки или твердый агар, дали обширные и ценные знания о поведении бактерий и механизмах толерантности к антибиотикам, эти результаты не всегда переводятся in vivo. Многие модели in vitro имеют ограниченную способность имитировать среду роста места инфекции человека, что требует дорогостоящих исследований in vivo. В свою очередь, многим моделям in vivo не хватает гибкости и разрешения, предоставляемых методами in vitro.
Синтетический муковисцидоз мокроты (SCFM2) предназначен для обеспечения среды для роста Па, аналогичной той, которая наблюдается во время хронической инфекции в легких муковисцидоза. SCFM2 включает источники питания, идентифицированные в отхаркиваемой смуте CF в дополнение к муцину, липидам и ДНК. Рост Pa в SCFM2 требует почти идентичного набора генов, необходимых для роста фактической мокроты, и поддерживает естественное образование агрегата Pa 9,10. После инокуляции планктонные клетки образуют агрегаты, которые увеличиваются в размерах за счет расширения. Отдельные клетки (называемые мигрантами) высвобождаются из агрегатов, мигрируют в неколонизированные районы и образуют новые агрегаты10. Эту историю жизни можно наблюдать с помощью CLSM и анализа изображений с разрешением одной клетки. Агрегаты Па, образующиеся в SCFM2, имеют размеры, аналогичные тем, которые наблюдаются в легкомCF 10. Эта модель позволяет наблюдать несколько агрегатов различного размера в режиме реального времени и в трех измерениях в микроновом масштабе. Покадровая микроскопия позволяет отслеживать тысячи (~50 000) агрегатов в одном эксперименте. Использование программного обеспечения для анализа изображений позволяет количественно оценить совокупные фенотипы по микроснимкам, включая совокупный объем, площадь поверхности и положение в трех измерениях до ближайших 0,1 мкм, как на индивидуальном агрегированном, так и на популяционном уровнях. Наличие способности группировать агрегаты по фенотипу и положению позволяет с точностью дифференцировать агрегаты на разных стадиях развития, а также их реакцию на изменениемикросреды 6,11.
Применение SCFM2 для изучения агрегатов Pa в анализах с низким объемом и высокой пропускной способностью делает его гибкой и экономически эффективной моделью. В качестве определенной среды SCFM2 обеспечивает однородность и воспроизводимость на нескольких платформах, обеспечивая питательную и физически релевантную методику изучения агрегатов Pa in vitro9. Приложения включают его использование в сочетании с CLSM для наблюдения пространственной организации и толерантности к антибиотикам при высоком разрешении (как описано в этой статье о методах). Возможность проводить эксперименты, которые предоставляют данные в микроновом масштабе в режиме реального времени, позволяет изучать внутривидовые и межвидовые взаимодействия, поскольку они могут происходить in vivo. Например, SCFM2 ранее использовался для изучения пространственной динамики клеточно-клеточной коммуникации в совокупных популяциях через сеть систем, используемых Pa для регулирования нескольких генов, которые способствуют вирулентности и патогенезу6.
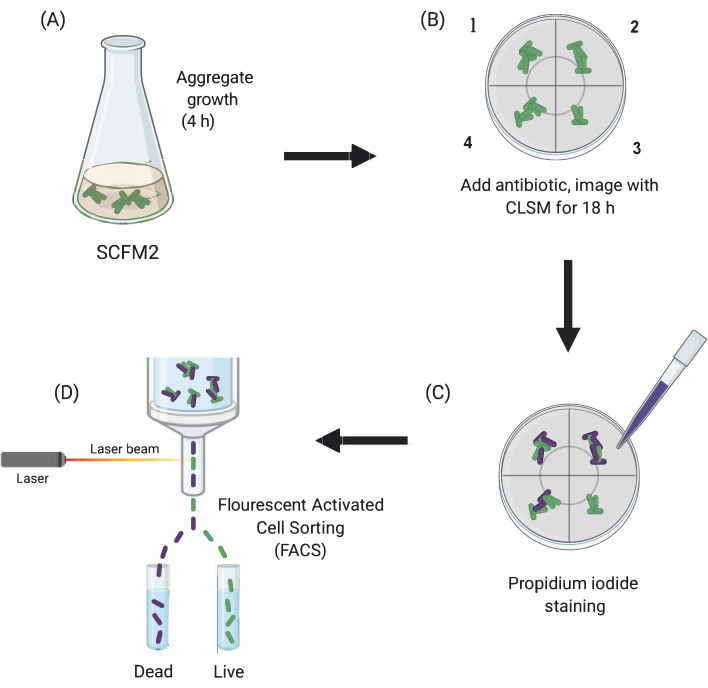
Рисунок 1:Графическое изображение основных экспериментальных этапов. (A)SCFM2 инокулируют Па-клетками и позволяют формировать агрегаты в чашке со стеклянным дном. (B) Агрегаты переносятся в конфокальный микроскоп и добавляют антибиотик. Изображены три технические реплики (камеры 1-3) и контрольная скважина (4) инокулированного SCFM2 без лечения антибиотиками. Агрегаты визуализируются с использованием CLSM в течение 18 ч. (C) После первоначальной 18-ч визуализации агрегаты обрабатывают йодидом пропидия для визуализации мертвых клеток и визуализируют с помощью CLSM (D) Агрегаты с желаемым фенотипом отделяют от SCFM2 с помощью FACS. Сокращения: SCFM2 = синтетический муковисцидоз мокроты; Pa = Синегнойная палочка; CLSM = конфокальная лазерная сканирующая микроскопия; FACS = флуоресцентная сортировка клеток. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Здесь демонстрируется полезность SCFM2 для изучения влияния лечения антибиотиками на агрегаты Pa в режиме реального времени, за которым следует использование клеточного подхода для выделения популяций агрегатов с различными фенотипами для последующего анализа(рисунок 1).
| Amino acids | |||
| Alanine | Acr s Organics s Organics |
56-41-7 | |
| Arginine HCl | MP | 1119-34-2 | |
| Asparagine | Acrs Organics |
56-84-8 | Prepared in 0.5 M NaOH |
| Cystine HCl | Alfa Aesar | L06328 | |
| Glutamic acid HCl | Acrs Organics |
138-15-8 | |
| Glycine | Acrs Organics |
56-40-6 | |
| Histidine HCl H2O | Alfa Aesar | A17627 | |
| Isoleucine | Acrs Organics |
73-32-5 | |
| Leucine | Alfa Aesar | A12311 | |
| Lysine HCl | Alfa Aesar | J62099 | |
| Methionine | Acrs Organics |
63-68-3 | |
| Ornithine HCl | Alfa Aesar | A12111 | |
| Phenylalanine | Acrs Organics |
63-91-2 | |
| Proline | Alfa Aesar | A10199 | |
| Serine | Alfa Aesar | A11179 | |
| Threonine | Acrs Organics |
72-19-5 | |
| Tryptophan | Acrs Organics |
73-22-3 | Prepared in 0.2 M NaOH |
| Tyrosine | Alfa Aesar | A11141 | Prepared in 1.0 M NaOH |
| Valine | Acrs Organics |
72-18-4 | |
| Antibiotic | |||
| Carbenicillin | Alfa Aesar | J6194903 | |
| Day-of Stocks | |||
| CaCl2 * 2H2O | Fisher Chemical | C79-500 | |
| Dextrose (D-glucose) | Fisher Chemical | 50-99-7 | |
| 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC) | Fisher (Avanti Polar Lipids) | 4235-95-4 | shake 15-20 min at 37 °C to evaporate chloroform |
| FeSO4 * 7H2O | Acrs Organics |
7782-63-0 | this stock equals 1 mg/mL, MUST make fresh |
| L-lactic acid | Alfa Aesar | L13242 | pH stock to 7 with NaOH |
| MgCl2 * 6H2O | Acrs Organics |
7791-18-6 | |
| N-acetylglucosamine | TCI | A0092 | |
| Prepared solids | |||
| Porcine mucin | Sigma | M1778-100G | UV-sterilize |
| Salmon sperm DNA | Invitrogen | 15632-011 | |
| Stain | |||
| Propidium iodide | Alfa Aesar | J66764MC | |
| Salts | |||
| K2SO4 | Alfa Aesar | A13975 | |
| KCl | Alfa Aesar | J64189 | add solid directly to buffered base |
| KNO3 | Acrs Organics |
7757-79-1 | |
| MOPS | Alfa Aesar | A12914 | add solid directly to buffered base |
| NaCl | Fisher Chemical | S271-500 | add solid directly to buffered base |
| Na2HPO4 | RPI | S23100-500.0 | |
| NaH2PO4 | RPI | S23120-500.0 | |
| NH4Cl | Acrs Organics |
12125-02-9 | add solid directly to buffered base |
| Consumables | |||
| Conical tubes (15 mL) | Olympus plastics | 28-101 | |
| Conical tubes (50 mL) | Olympus plastics | 28-106 | |
| Culture tubes w/air flow cap | Olympus plastics | 21-129 | |
| 35 mm four chamber glass-bottom dish | CellVis | NC0600518 | |
| Luria Bertani (LB) broth | Genessee Scientific | 11-118 | |
| Phosphate-buffered saline (PBS) | Fisher Bioreagents | BP2944100 | |
| Pipet tips (p200) | Olympus plastics | 23-150RL | |
| Pipet tips (p1000) | Olympus plastics | 23-165RL | |
| Serological pipets (5 mL) | Olympus plastics | 12-102 | |
| Serological pipets (25 mL) | Olympus plastics | 12-106 | |
| Serological pipets (50 mL) | Olympus plastics | 12-107 | |
| Ultrapure water (RNAse/DNAse free); nanopure water | Genessee Scientific | 18-194 | Nanopure water used for preparation of solutions in Table 1 |
| Syringes (10 mL) | BD | 794412 | |
| Syringes (50 mL) | BD | 309653 | |
| 0.22 mm PES syringe filter | Olympus plastics | 25-244 | |
| PS cuvette semi-mico | Olympus plastics | 91-408 | |
| Software | |||
| Biorender | To prepare the figures | ||
| FacsDiva6.1.3 | Becton Dickinson, San Jose, CA | ||
| Imaris | Bitplane | version 9.6 | |
| Zen Black | |||
| Equipment | |||
| FacsAriallu | Becton Dickinson, San Jose, CA | ||
| LSM 880 confocal laser scanning microscope | Zeiss |
