Intracellular intake and extracellular glucose depletion were measured in 3T3-L1 preadipocytes, in response to different concentrations of FD-glucose (Figure 2) with and without insulin stimulation. Figure 2A demonstrates a dose-dependent increase in the intracellular uptake of FD-glucose, which was significantly increased in the presence of insulin. The concomitant decrease in extracellular FD-glucose in the same cells is shown in Figure 2B, where insulin stimulation led to significantly decreased extracellular FD-glucose levels compared to the samples without insulin stimulation. The intracellular intake of FD-glucose correlated with the extracellular depletion of FD-glucose in a dose-dependent manner in samples with and without insulin (Figure 2C). Thus, extracellular depletion of FD-glucose can measure a change in glucose uptake with comparable accuracy (R2 = 0.99, P < 0.001) as intracellular FD-glucose uptake.
Next, an experimental setting was developed to validate extracellular FD-glucose uptake application in kinetic studies examining different tissues upon stimulation with canonic and experimental inducers of glucose uptake (Figure 3). We selected the Lepob mouse model of insulin resistance in peripheral tissues, including visceral white adipose tissue27,28. The well-established responses to insulin in these mice were compared with effects of novel compound AAC2, acting on both peripheral and nervous tissues via leptin receptor and glucose transporter GLUT1 dependent mechanisms18. Lepob mice were i.p. injected with insulin or AAC2. Organs were dissected 15 min after injection. Then, the kinetics of extracellular FD-glucose depletion was studied in visceral fat, liver, and brain ex vivo (Figure 3). In agreement with the reported insulin resistance in peripheral tissues, extracellular FD-glucose was not depleted in non-stimulated control visceral fat during 120 min of incubation (Figure 3A). In contrast, pretreatment of mice with insulin or AAC2 before the dissection led to significant depletion of extracellular FD-glucose within a time interval of 30 min and 60 min, respectively.
Liver uptake of glucose utilizes glucose transporters, including GLUT2, that is independent of insulin stimulation29,30. Accordingly, extracellular FD-glucose was initially not depleted in insulin-stimulated liver explants compared with non-stimulated liver explants (Figure 3B), although significant depletion of FD-glucose was observed after 120 min of incubation in liver explant stimulated with insulin compared with the initial levels (0 min). On the other hand, extracellular FD glucose was significantly decreased in a time-dependent manner in liver explants pretreated with AAC2.
In the brain, the main transporter is GLUT1 among other transporters31. The FD-glucose depletion kinetics was strikingly different in the brain compared to other tissue (Figure 3C). The significant depletion of glucose was observed in the extracellular medium containing the non-treated brain after 60 min of incubation (from 100% at 0 min to 78%). The medium incubated with brains from insulin-treated Lepob mice have shown moderate but a significant linear decrease in FD-glucose (from 100% at 0 min to 95%). AAC2-stimulated brains lead to a profound rapid decrease of extracellular FD-glucose during the first 20 min (from 100% at 0 min to 67.4%). Together, measurements of extracellular glucose depletion support the previously reported differences in glucose uptake in these tissues in vivo32.
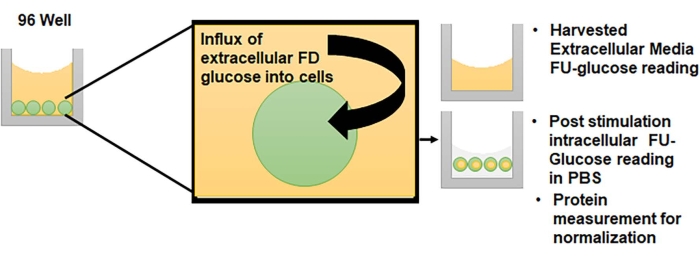
Figure 1: Principle of the extracellular FD-glucose depletions method. Cells cultured in 96-well format are suitable for this assay. To survive, cells take up glucose and this influx decreases the levels of extracellular glucose. The replacement of glucose with FD-glucose allows monitoring of these changes. Please click here to view a larger version of this figure.
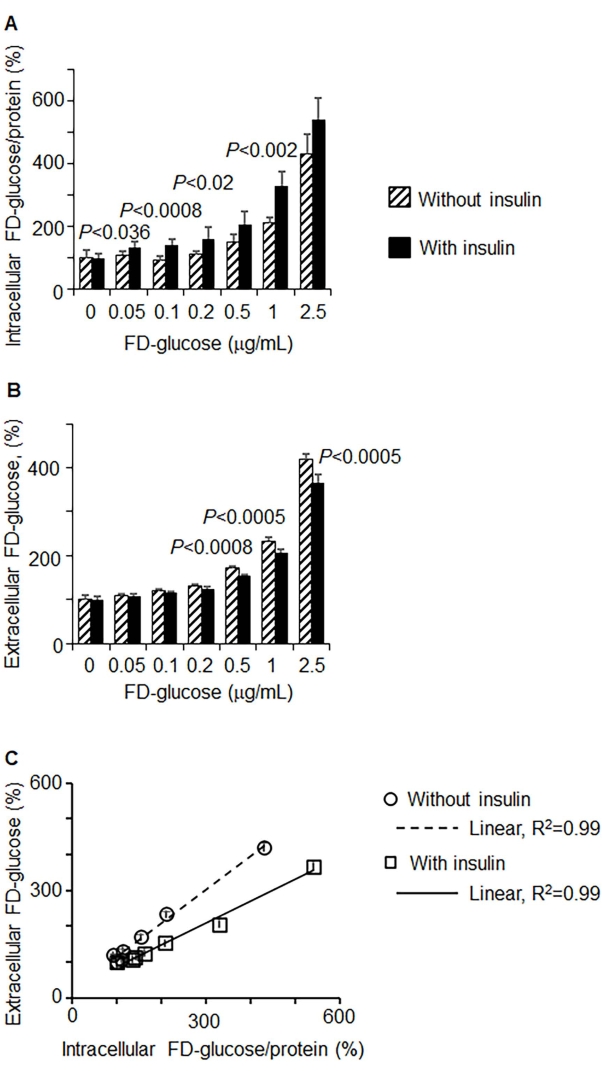
Figure 2: Depletion of extracellular FD-glucose correlated with the intracellular uptake in vitro. 3T3-L1 preadipocytes were incubated in a glucose-free medium for 40 min before incubation with different concentrations of FD-glucose with and without insulin (1.7 mM; 40 min incubation). (A, B) Dose-dependent uptake of FD-glucose (A) and extracellular FD-glucose depletion (B) in control (i.e., without insulin) (hatched bars) and insulin-stimulated cells (black bars). FD-glucose uptake was normalized by protein concentrations. Data are shown as % of the value measured in control incubated without FD-glucose (mean SD, n = 8 per condition). Significance was examined by unpaired t-test. (C) Correlation between intracellular and extracellular FD-glucose (%) measures with (squares) or without (circles) insulin in experiments described in (A) and (B). Pearson correlation, P < 0.001. Please click here to view a larger version of this figure.
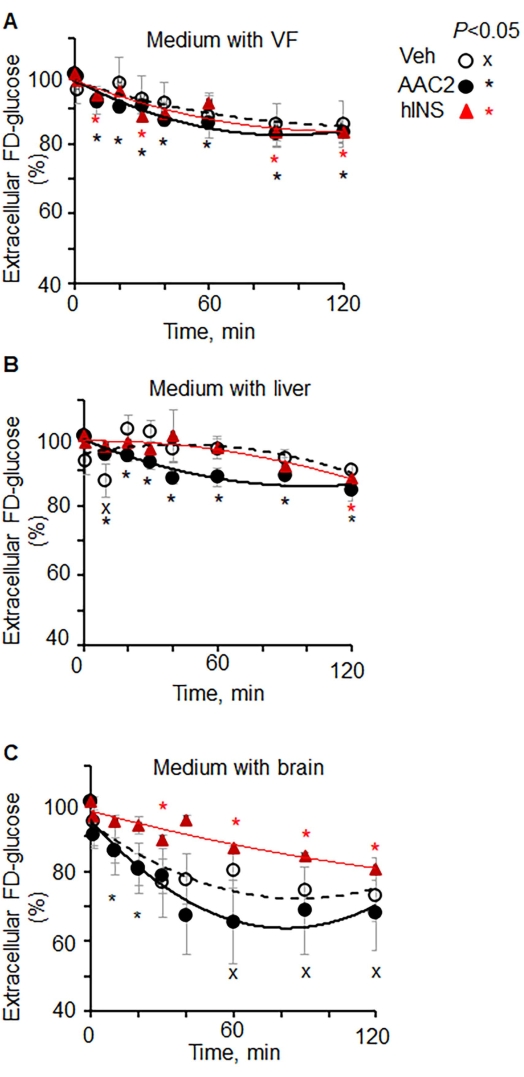
Figure 3: Kinetics of extracellular FD-glucose depletion in different organs ex vivo. (A–C) Lepob mice were injected with vehicle (PBS, n = 4), insulin (12 IU/kg BW, n = 3), and AAC2 (1 nmol/g BW, n = 3). After 15 min, tissues were dissected and isolated. Explants of (A) visceral fat, (B) liver, and (C) brain were incubated in FD-glucose (0.29 mM). The kinetics of extracellular glucose depletion was measured in aliquots of the medium at different time points. Data (mean SD) are shown as % of fluorescence in each organ at 0 min of incubation. Significance P < 0.05, unpaired t-test. Please click here to view a larger version of this figure.
| Solution/Medium | Components |
| Ethanol:DMSO (1:1/v/v) | Ethanol (200 µL) and DMSO (200 µL) in 1-1.5 mL tube; use cell culture grade ethanol and DMSO |
| Medium 1 | DMEM (89 mL), Penicillin/streptomycin (1%) (1 mL) and Calf serum (10%) (10 mL) in sterile 50 mL tube |
| Centrifugation Medium 2 | DMEM (89 mL), Penicillin/streptomycin (1%) (1 mL) and Bovine serum (10%) (10 mL) in sterile 50 mL tube |
| Storage FD-glucose solution 5 mg/mL (14.5 mM) | FD glucose (1 mg) and Ethanol:DMSO (1:1/v/v) (200 µL) in 0.5 mL tube; store at -80 °C, preferentially under argon or nitrogen atmosphere |
| Working FD-glucose solution 5 µg/mL (14.5 mM) | Storage FD glucose solution 5 mg/mL (1 µL), Glucose free DMEM (999 µL); prepare immediately before experiment. |
Table 1: Preparation of culture media and FD-glucose solutions.
| Dilution | Control | 1:2×103 | 1:5×103 | 1:1×104 | 1:2.5×104 | 1:5×104 | 1:1×105 |
| Concentration (µg/mL) | 0 | 2.5 | 1 | 0.5 | 0.2 | 0.1 | 0.05 |
| Glucose-free DMEM (µL) | 1000 | 500 | 800 | 900 | 960 | 980 | 990 |
| FD Glucose 5 µg/mL (µL) | 0 | 500 | 200 | 100 | 40 | 20 | 10 |
Table 2: Preparation of FD-glucose solutions with different concentrations for experiments shown in Figure 2.
