The co-culture system described in Figure 1 allows for a controlled study of HIV-1 innate sensing. Due to the variable nature of primary cells, it is important that normal ratios of myeloid and T cells (CD14+ and CD3+ respectively) are observed, such as in the example shown in Figure 2A. Furthermore, only a low number of activated T cells (higher FS and higher CD3 levels as compared to non-activated T cells) are desirable in the freshly isolated PBMCs cultures. PBMCs with abnormal ratios between the various cell-types are often incapable of mounting a proper innate immune response in vitro, likely due to an already activated phenotype as a consequence of an ongoing infection. Most importantly, the relative percentage of pDCs needs to be evaluated as shown in Figure 2B. Although this percentage can vary from 0.2 to 1.2%, an abnormal sensing phenotype is also observed with both extremes of this range. Enrichment of pDCs using negative selection often yields 50-95% pDC purity, as seen in the two examples from in Figure 2B (92% and 72%). A prototypical example for the overall experiment is shown in Figures 3 and 4. In this example, MT4 cells were infected with pNL4.3-GFP_ires_Nef WT or delta Vpu to achieve 30% infection at the time of co-culture (Figure 3A). As previously described, only the infections with WT virus resulted in a significant down-regulation of surface BST2 (Figure 3B). Due to their differences in size and granularity, MT4 cells can be easily distinguished from PBMCs by flow cytometry (Figure 4A). After the co-culture incubation, only PBMCs in contact with known TLR7 or TLR9 agonist and PBMCs in contact with HIV-1 infected cells released significant amounts of type-I IFN (Figure 4B). No IFN was detected in PBMCs alone, in PBMCs co-cultured with mock-infected MT4 cells, or in infected MT4 cells alone (Figure 4B). In the example presented here, innate sensing of HIV-1-infected MT4 cells by PBMCs was found to be significantly reduced in the presence of Vpu.
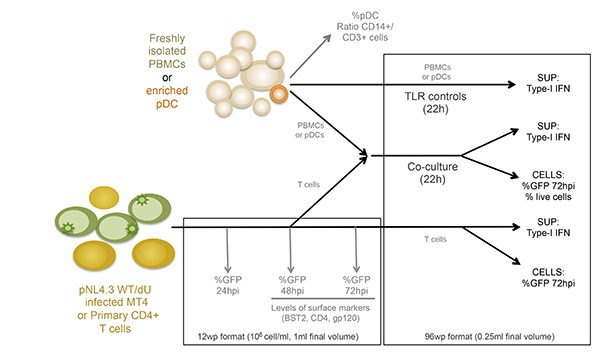
Figure 1: Schematic overview of the experimental design. SUP: supernatant Click here to view larger image.
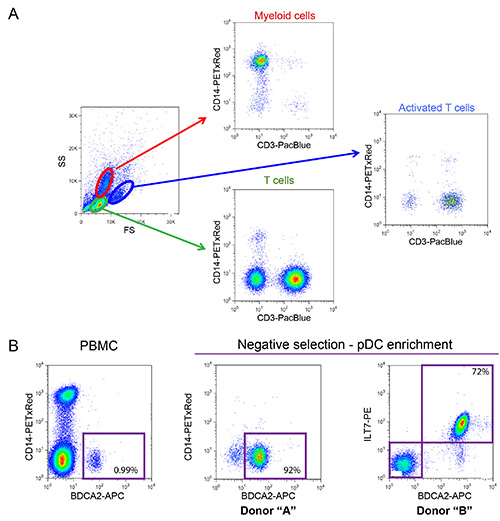
Figure 2: Phenotypic characterization of freshly isolated PBMCs and enriched pDCs. (A) Normal distribution of myeloid cells and T cells observed in PBMCs isolated from a healthy donor (Percentage of myeloid cells should range between 10-20% and T cells between 55-65%). PBMCs sample were stained using fluorophore-conjugated antibodies that recognize surface CD14 (myeloid lineage marker) and CD3 (T cell marker). (B) Phenotypic characterization of pDCs. Prior to enrichment pDCs represent between 0.2-1.2% of the total number of cells within the PBMCs (0.99% in the example shown). These cells can be identified based on expression of specific markers, such as BDCA2 and ILT7, present at the cell surface. Negative selection can be used to significantly enrich the number of pDCs and eliminate potentially detrimental contaminations, such as CD14+ myeloid cells (enrichment reaching 92% and 72% in the two examples shown). Click here to view larger image.
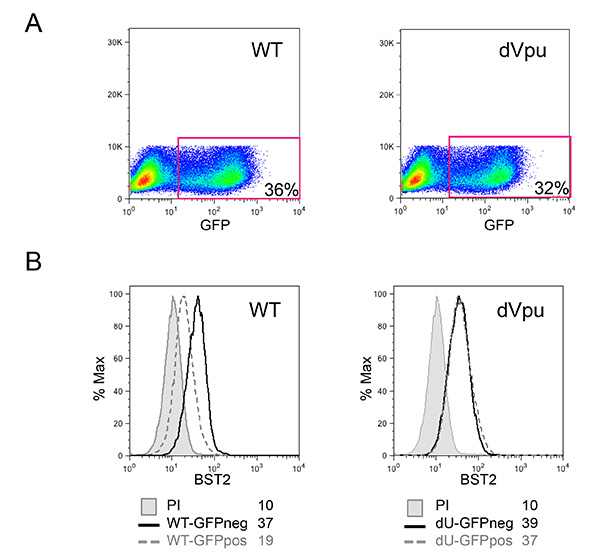
Figure 3: Characterization of HIV-1-infected target MT4 cells. (A) Percentage of infected cells within the MT4 cultures infected with pNL4.3-GFP_ires_Nef WT or dVpu as determined by the percentage of GFP positive cells. (B) BST2 cell surface expression in the infected cells was evaluated after surface staining. The grey filled histograms represent mock-infected cells stained with the pre-immune rabbit serum (unstained control); the remaining histograms represent cells stained with anti-BST2 polyclonal rabbit serum. Histograms with solid lines represent control GFP negative (neg) cells; while histograms with dotted lines correspond to infected GFP positive (pos) cells. Mean fluorescence intensity (MFI) values are indicated for each sample. Flow cytometry was performed and analyzed as described in Figure 2. Click here to view larger image.
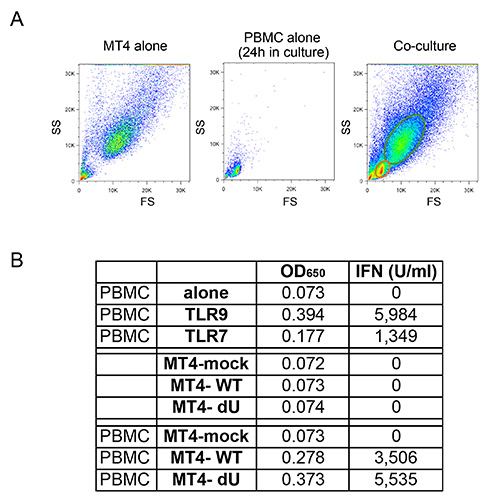
Figure 4: Co culture of freshly isolated PBMCs and infected MT4 T cells. (A) Comparison of flow cytometry FS/SS profiles observed for MT4 T cells alone, PBMCs alone, or co-cultures between PBMCs and MT4 T cells. Due to differences in size and granularity, MT4 T cells can be easily distinguished from PBMCs in co-cultures using flow cytometry. (B) Representative example of the amount of type-I IFN released after stimulation of PBMCs with TLR7 or TLR9 agonist (ago), or after co-cultures with mock or the indicated MT4 cells infected with pNL4.3-GFP_ires_Nef WT or dVpu. Raw data shown as OD650 values and their corresponding converseion to type-I IFN concentration expressed in U/ml. Click here to view larger image.
