Using Microtiter Dish Radiolabeling for Multiple In Vivo Measurements Of Escherichia coli (p)ppGpp Followed by Thin Layer Chromatography
Summary
The growth of radiolabeled bacterial cultures in microtiter dishes facilitates high throughput sampling that allows multiple technical and biological replicate assays of nucleotide pool abundance, including that of (p)ppGpp. The effects of growth transitions provoked by sources of physiological stress as well as recovery from stress can be monitored.
Abstract
The (p)ppGpp nucleotide functions as a global regulator in bacteria in response to a variety of physical and nutritional stress. It has a rapid onset, in seconds, which leads to accumulation of levels that approach or exceed those of GTP pools. Stress reversal occasions a rapid disappearance of (p)ppGpp, often with a half-life of less than a minute. The presence of (p)ppGpp results in alterations of cellular gene expression and metabolism that counter the damaging effects of stress. Gram-negative and Gram-positive bacteria have different response mechanisms, but both depend on (p)ppGpp concentration. In any event, there is a need to simultaneously monitor many radiolabeled bacterial cultures at time intervals that may vary from 10 seconds to hours during critical stress transition periods. This protocol addresses this technical challenge. The method takes advantage of temperature- and shaker-controlled microtiter dish incubators that allow parallel monitoring of growth (absorbance) and rapid sampling of uniformly phosphate-radiolabeled cultures to resolve and quantitate nucleotide pools by thin-layer chromatography on PEI-cellulose. Small amounts of sample are needed for multiple technical and biological replicates of analyses. Complex growth transitions, such as diauxic growth and rapid (p)ppGpp turnover rates can be quantitatively assessed by this method.
Introduction
The (p)ppGpp second messenger is a global regulator that modulates the expression of a large number of genes, including genes for synthesizing ribosomes and amino acids1,2. Although initially discovered in Escherichia coli3, (p)ppGpp can be found in both Gram positive and Gram negative bacteria as well as in plant chloroplasts4,5. For E. coli and other Gram-negative bacteria, (p)ppGpp interacts directly with RNA polymerase at two different sites6,7,8. In Gram positives, (p)ppGpp inhibits GTP abundance, which is sensed by CodY, a GTP-binding protein with gene-specific DNA recognition motifs that lead to regulation9,10. (p)ppGpp accumulates in response to starvation for different nutrients and stress conditions, resulting in slow growth and adjustments of gene expression to allow adaptation to stress11,12.
The net amount of ppGpp accumulated reflects a balance between synthetase and hydrolase activities. In E. coli RelA is a strong synthetase and SpoT is bifunctional, with a strong hydrolase and a weak synthetase, each of which might be regulated differently in a stress dependent manner. The strong RelA synthetase is activated when the availability of codon-specified charged tRNA bound to the ribosomal A site when it fails to keep up with the demands of protein synthesis13,14,15. The weak SpoT (p)ppGpp synthetase is activated while the strong (p)ppGpp hydrolase is inhibited in response to other stress conditions and through other mechanisms. Under some conditions, proteins such as ACP or Rsd can bind to SpoT, which also change the balance between hydrolysis and synthesis16,17. In Gram positives, synthesis and hydrolysis reflect a more complex balance between a single RelA SpoT homolog (RSH) protein with strong synthesis and hydrolysis activities as well as smaller hydrolases and/or synthetases12.
The (p)ppGpp nucleotides were first discovered as unusual 32P labeled spots that appeared on autoradiograms of thin-layer chromatograms (TLC) during a stringent response induced by amino acid starvation3. More detailed labeling protocols have been reviewed 18. The protocol described here (Figure 1) is a modification of these protocols that allows monitoring growth of multiple samples on microtiter plates. This facilitates multiple biological and technical estimates of (p)ppGpp abundance changes and was initially developed for studies of diauxic shifts19. Labelling of (p)ppGpp with 32P and detection by TLC also allows measurements of (p)ppGpp degradation rates. Alternative methods have been developed to determine (p)ppGpp levels such as mass spectrometry, HPLC20, fluorescent chemosensors21,22, and GFP gene fusions to promoters affected by ppGpp23,24. Fluorescent chemosensors currently have a limited use due small spectral shifts after binding ppGpp as well as problems distinguishing between ppGpp and pppGpp21. This method is efficient to detect (p)ppGpp in vitro, but not in cellular extracts. Methods involving HPLC have been improved20 but require expensive equipment and are not well adapted to high through-put. Finally, GFP fusions can give an estimate of ppGpp dependent activation or inhibition, but do not measure ppGpp itself. While each of these alternative methods are valuable, they require expensive equipment or substantial hands-on time, or they are otherwise not amenable to multiple kinetic sampling and subsequent processing. With the method described here, 96 samples can be applied to six TLC plates in about 20 min (18 samples per plate), resolved by TLC development in less than a couple of hours, with quantitative data obtained after several hours or overnight, depending on labeling intensity.
Protocol
1. Media Preparation
- For MOPS (3-(N-morpholino)propanesulfonic acid) media25, use 1/10 volume of 10x MOPS salts, 1/100 volume of 100x micronutrients solution, 3 mM sodium phosphate for overnight cultures or 0.2 mM sodium phosphate for uniform labeling with 32P, 0.2% glucose and 1 µg/mL thiamin (vitamin B1). Add amino acids at 40 µg/mL if required.
- For MOPS salts, use 400 mM MOPS, 40 mM Tricine, 0.1 mM FeSO4, 95 mM NH4Cl, 2.6 mM K2SO4, 5 µM CaCl2, 10.56 mM MgCl2, and 500 mM NaCl. Add the different components in the order written to avoid precipitation. Adjust to pH 7.4 with KOH and sterilize by filtration.
- For 100x micronutrients solution, use 3 µM (NH4)6(MO7)24, 0.4 mM H3BO4, 30 µM CoCl2, 10 µM CuSO4, 80 µM MnCl2, and 10 µM ZnSO4. Sterilize by autoclaving.
- For glucose (20%) and sodium phosphate (300 mM) solutions, sterilize as separate 100x stocks by autoclaving. Sterilize 100x stocks of thiamine and amino acid mixtures by filtration. Prepare fresh media from sterile stock solutions for each experiment.
2. Labelling with 32P
- Grow overnight cultures in MOPS media with 3 mM sodium phosphate.
- In a 24 well plate, add 550 µL of MOPS media containing 0.2 mM sodium phosphate carrier and 30 µL of each bacterial inoculum (dilution 1/20). This will produce an initial inoculum of OD600nm 0.1. Use one well with fresh media as a sterility control.
- Place the plate on a shaking incubator at 37 °C shaking at 900 rpm for 30 min. Cultures are heated from below and shaking provides aeration. Attach the plate firmly with tape to avoid tilting or spills. Growth can be monitored with a replicate plate in parallel in media lacking 32P using a plate reader and incubating under the same conditions.
- Add 100 µCi to each well of 32P orthophosphate (20 µL from a 5 mCi/mL stock).
NOTE: The amount of radioactivity can also be adjusted to meet experimental needs. For low basal levels 100 µCi with 0.2 mM carrier phosphate works well, but for measuring (p)ppGpp levels that are expected to become equivalent to GTP pools, label can be dropped to 25 or 50 µCi. Lowering carrier phosphate below 0.2 mM is not recommended to avoid phosphate levels becoming limiting for growth.
CAUTION: 32P is a β emitting isotope so use proper shielding specified for safety. All radioactive material must be properly disposed taking all the possible precautions. - Grow in shaking incubator for 1 to 2 doublings (1 h or more) to allow external label to largely equilibrate with intracellular nucleotide pools before inducing stress.
3. Induction of Stress or Starvation
- Induce stress using a method of your choosing, depending on the sort of stress studied, e.g. adding metabolic pathway inhibitors, changing temperature, exhausting required nutrients, shifting osmolarity shifts, inducing oxidative stress, adding toxins, etc.
- For example, add 100 µg/mL L-valine to induce isoleucine starvation in E. coli K-12 strains. Increased levels of ppGpp will be observed after 5 minutes of induction.
4. Sampling and ppGpp Extraction
- Add 20 µL of each labelled cell sample to a 0.2 mL PCR tube containing 20 µL of ice cold 6 M Formic acid.
- Immediately place the samples in dry ice.
- Enhance cellular extraction efficiency by 3 cycles of freezing and thawing.
- Just before spotting PEI cellulose thin layer chromatograms, centrifuge samples for 1 min at maximum speed to pellet cell debris to avoid spotting it on chromatograms.
5. Thin-layer Chromatography
- With a soft pencil, mark an origin line 1 cm from the edge of the 20 cm x 20 cm PEI-Cellulose TLC plate that has had 5 cm removed from the top with scissors. Apply 5 µL as a droplet in the PEI surface for each sample. Begin ascending development without allowing this spot to dry, which improve resolution by minimizing streaking.
- Do ascending development in a tank with a layer of 1.5 M KH2PO4 (pH 3.4) solution shallow enough so that liquid does not touch the origin sample spot. Cover the tank with an air-tight seal and allow liquid ascent to the top of the trimmed sheet, 15 cm. To achieve pH 3.4 for a 1.5 M KH2PO4 solution, it is necessary adjust the pH by addition of H3PO4.
- Remove the fully developed chromatogram and air dry at room temperature.
- Cut and discard the top (pH front) portion of the chromatogram containing the free 32P into radioactive waste. This portion is represented in yellow color in Figure 2 and easily visualized under UV light. If measuring only (p)ppGpp nucleotides that migrate slower than GTP, it is recommended to run a UV-visible GTP standard and then cut and discard a larger upper portion (anything above GTP, as shown in Figure 2).
- Expose autoradiographic films overnight with a phosphor screen.
- Develop the film. Capture and quantitate the phosphor screen signal with a phosphoimager.
6. Quantitation of (p)ppGpp
- Quantitate radioactive spots with Image J26,27.
NOTE: The amount of (p)ppGpp can be normalized to total amount of G nucleotides observed in each sample (Figure 2). If the amount of background is homogeneous, a single blank (box 4 from Figure 2) can be subtracted to correct for background. Total G is the sum of GTP + ppGpp + pppGpp detected. This normalization assumes that GMP and GDP levels are negligible, which is true for E. coli. The ratio of a given nucleotide to total G provides an important way to correct for variations of applied sample volume.
7. Measurement of the Rate of ppGpp Decay
NOTE: A modification of this protocol allows measurements of ppGpp decay rates.
- After provoking stress or starvation for a time sufficient to allow ppGpp accumulation (step 3), reverse the stress in order to block continued ppGpp synthesis, which allows detection of hydrolytic rates. The procedure for reversal will depend on the stress. For example, add 200 µg/mL of chloramphenicol to reverse any amino acid starvation.
- Decay rates are usually rapid but may vary, so take samples every 20-30 s up to 2 min. Process samples as in step 4.
- Once the TLC is developed (as in step 5) and the levels of ppGpp obtained during the time course, plot residual ppGpp content on a semi-logarithmic plot vs time to allow visualization of zero order rates of decay, as a straight line, and estimation of the ppGpp half-life.
Representative Results
In E. coli K-12 strains, the addition of valine provokes an endogenous starvation of isoleucine, which results an increase of ppGpp levels after 5 min3. Cells grown in MOPS containing all amino acids except for ILV were labeled with 32P as indicated in Figure 1. Once labelled, 6 µL of 10 mg/mL L-valine (100 µg/mL final concentration) was added to produce isoleucine starvation. Samples were taken 0 and 5 min after the addition of valine. After 5 min (Figure 3A), a 2 and 2.5-fold increase in the levels of ppGpp and pppGpp occured. As a negative control, a cell-free labelled sample was used to detect possible compounds that were not orthophosphate in the 32P source. Also, including a ppGpp deficient strain (ΔrelA ΔspoT) as a negative control could help a better identification of the spots.
Reversal of isoleucine starvation can be achieved by chloramphenicol, an inhibitor of protein synthesis, which reduces consumption of charged ile-tRNA and in turn, restores high ratios of charged to uncharged tRNA. Activation of the strong RelA-mediated ppGpp synthetase is abolished, which allows a measure of ppGpp degradation unperturbed by residual synthesis. Therefore, 200 µg/mL of chloramphenicol was added to the starved cultures and samples were taken at 20 s intervals thereafter. In this case, ppGpp decayed with a half-life of about 64 s (Figure 3B).
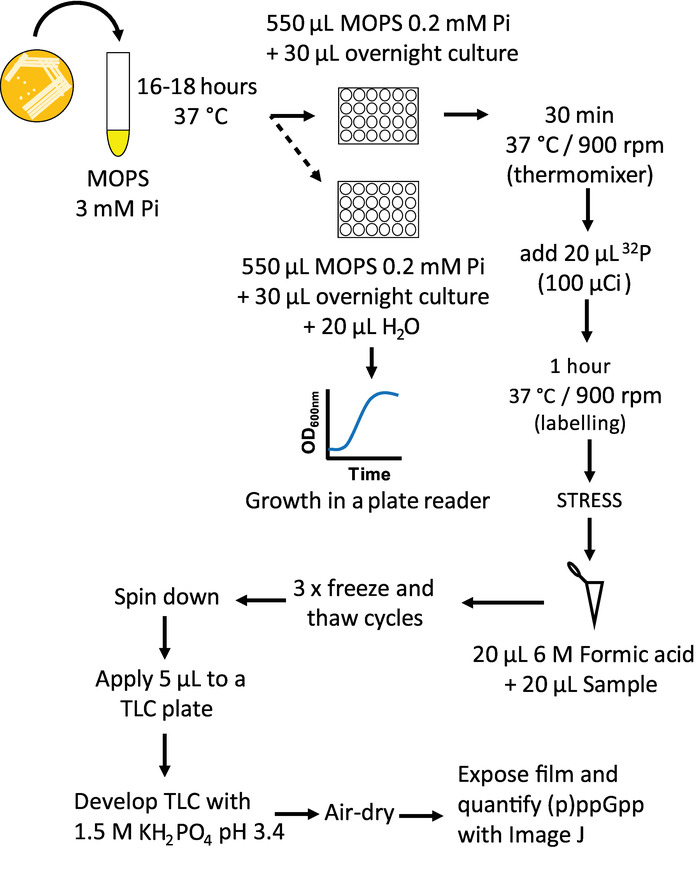
Figure 1: Measurement of ppGpp by TLC workflow. Schematic representation of the protocol to extract ppGpp form bacterial cultures and its posterior detection by TLC. Please click here to view a larger version of this figure.
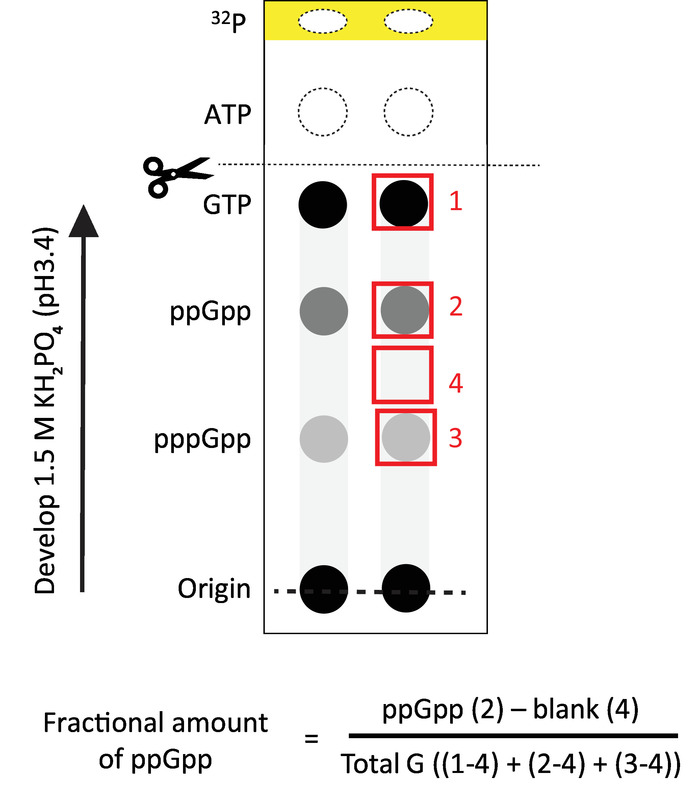
Figure 2: Representative TLC analysis. Schematic representation of the results obtained after autoradiography and phosphor screen overnight with the TLC. The spots to be measured are shown in red. The formula to calculate the fractional amount of ppGpp is also shown. Total G refers to the total amount of GTP + ppGpp + pppGpp detected in the sample. The yellow band represent the pH front visible under UV light. Scissors indicate where we recommend cutting the chromatogram for discarding most of the radioactivity. The arrow indicates the direction of the flow during ascending development with 1.5 M phosphate buffer. Please click here to view a larger version of this figure.
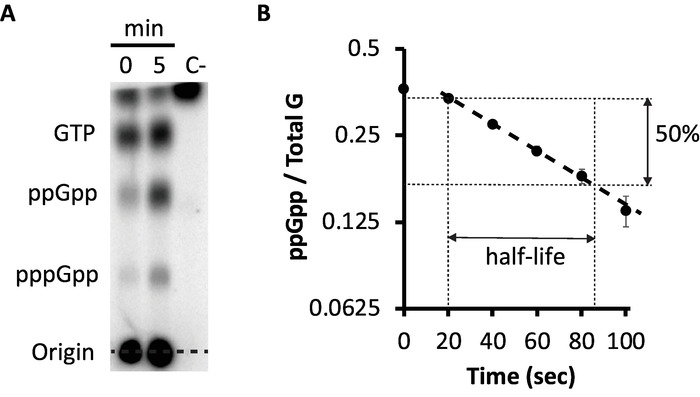
Figure 3: measurement of synthesis and decay of ppGpp. (A) Detection of ppGpp by TLC for samples 0 and 5 minutes after addition of valine. Spots corresponding to GTP, ppGpp and pppGpp are marked. A cell-free culture was included as negative control (C-). (B) Decay of ppGpp after addition of chloramphenicol to reverse amino acid starvation. The ppGpp half-life is determined from the exponential decay rate represented by dotted lines. Error bars reflect SD from duplicates. Y-axis is represented in a semilogaritmic (log2) scale. Please click here to view a larger version of this figure.
Discussion
Achieving near uniform labeling of the cells is a critical step for this protocol. Therefore, the use of defined media, such as MOPS or Tris media, is crucial to allow variation of carrier phosphate concentrations and specific activity. Phosphate buffered media, such as M9 or media A, cannot be used. Most undefined media contain variable amounts of phosphate, such as LB, tryptone and casamino acids. The phosphate isotope 33P is a weaker emitter that can be substituted for 32P. Advantages of the substitution are that it is safer and has a half-life of 25 days instead of 14 days for 32P. However, the more energetic emissions of 32P considerably enhance detection sensitivity. It is important to stress the dangers of working with isotopes and the importance of using proper shielding protections and disposal. The recent discovery of a riboswitch able to detect specifically ppGpp28 may someday lead to a safer way to detect ppGpp in vitro and in vivo. As mentioned in the introduction, other methods exist, although the use of phosphate labelling remains a sensitive, direct and rapid approach for through-put measurements of bacterial (p)ppGpp as well as other phosphate labeled compounds, such as ribo- or deoxyribo-NTP pools, PRPP, PPi or sugar phosphates.
Another critical step is to ensure that the growth of the parallel cultures (shaking incubator and plate reader) is similar. When studying nutrient exhaustion, such as diauxic shift19, the synchrony between cultures is crucial. The comparability of growth on labeled and unlabeled plates can be assessed by OD600 measurements of an unlabeled well on the otherwise radioactive plate. To increase the number of samples tested, a 96 well plate can be used instead 19, but the amount of media required to minimize evaporation will reduce culture aeration. Basal levels of (p)ppGpp will be slightly higher during growth in a 96 well plate compared to a 24 well plate due to the reduced aeration. Therefore, for measuring basal levels, a 24 well plate growth is recommended. To measure variations of more strongly induced (p)ppGpp levels, a 96 well plate is preferred. The choice between 24 and 96 well microtiter plates depends on the experimental goal.
Variations of this protocol have many potential applications for different conditions of stress. Recently we have applied it to measure accumulation and decay of ppGpp during classical diauxic shifts from glucose to lactose and to other alternative sugars. These studies revealed involvement of both glucose starvation and amino acid starvation19. Here we describe amino acid starvation induced by valine in E. coli K12 strains as a simpler and well-studied stress condition3. This example might be used as a control before performing more complicated starvation situations. Amino acid starvation can be provoked by adding limited amounts of a required amino acid that is exhausted during growth; adding an inhibitor of tRNA aminoacylation or synthesis (serine hydroxamate, mupirocin, or 3-aminotriazole) or for E. coli K12 adding valine in the absence of isoleucine). Serine hydroxamate is often used to inhibit tRNASer aminoacylation but requires high concentrations, which may cause problems due to the hydroxamate reactivity with other compounds. Reversal of serine hydroxamate effects by adding a large amounts of serine can have non-desired effects1. Although SHX has been proven to be effective as a diagnostic of ppGpp production, we do not recommend its use for physiological studies to produce amino acid starvation, because normal (p)ppGpp feedback mechanisms are abolished, such as physiological consequences of lowering superfluous activities but allowing activation of stress survival regulatory circuits.
Modifications of the TLC procedures described here are necessary in order to properly separate noncanonical regulatory nucleotides, such as (p)ppApp29, from mixed samples with (p)ppGpp. It has been shown that pppApp has an antagonistic role to ppGpp in vitro30,31; therefore, better separation of both compounds is required for future studies.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
This research was supported the Intramural Research Program, Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH.
Materials
| (NH4)6(MO7)24 | Fisher Scientifics | A-674 | |
| Autoradiography film | Denville scientific inc. | E3218 | |
| CaCl2 | J.T.Baker | 1-1309 | |
| Chloramphenicol | RPI | C61000-25.0 | |
| CoCl2 | Fisher Scientifics | C-371 | |
| CuSO4 | J.T.Baker | 1843 | |
| FeSO4 | Fisher Scientifics | I-146 | |
| Formic acid | Fisher Biotech | BP1215-500 | |
| Glucose | Macron | 4912-12 | |
| H3BO4 | Macron | 2549-04 | |
| H3PO4 | J.T.Baker | 0260-02 | |
| K2SO4 | Sigma | P9458-250G | |
| KH2PO4 | Fisher Biotech | BP362-500 | |
| L-Valine | Sigma | V-6504 | |
| MgCl2 | Fisher Scientifics | FL-06-0303 | |
| Microplate reader Synergy HT | Biotek | Synergy HT | |
| MnCl2 | Sigma | M-9522 | |
| MOPS | Sigma | M1254-1KG | |
| Na2HPO4 | Mallinckrodt | 7892 | |
| NaCl | J.T.Baker | 3624-01 | |
| NaH2PO4 | Mallinckrodt | 7917 | |
| NH4Cl | Sigma | A0171-500G | |
| P-32 radionuclide, orthophosphoric acid in 1 mL water (5 mCi) | Perkin Elmer | NEX053005MC | |
| Storage phosphor screen | Kodak | So230 | |
| Thermomixer | Eppendorf | 5382000015 | |
| Thiamine | Sigma | T-4625 | |
| TLC PEI Cellulose F | Merk-Millipore | 1.05579.0001 | |
| Tricine | RPI | T2400-500.0 | |
| Typhoon 9400 imager | GE Healthcare | ||
| ZnSO4 | Fisher Scientifics | Z-68 |
Riferimenti
- Cashel, M., Gentry, D., Hernandez, V., Vinella, D. The stringent Response. Escherichia coli and Salmonella: Cellular and Molecular Biology. , 1458-1489 (1996).
- Magnusson, L. U., Farewell, A., Nyström, T. ppGpp: a global regulator in Escherichia coli. Trends in Microbiology. 13 (5), 236-242 (2005).
- Cashel, M., Gallant, J. Two Compounds implicated in the Function of the RC Gene of Escherichia coli. Nature. 221 (5183), 838-841 (1969).
- Braeken, K., Moris, M., Daniels, R., Vanderleyden, J., Michiels, J. New horizons for (p)ppGpp in bacterial and plant physiology. Trends in Microbiology. 14 (1), 45-54 (2006).
- Atkinson, G. C., Tenson, T., Hauryliuk, V. The RelA/SpoT homolog (RSH) superfamily: distribution and functional evolution of ppGpp synthetases and hydrolases across the tree of life. PloS One. 6 (8), e23479 (2011).
- Mechold, U., Potrykus, K., Murphy, H., Murakami, K. S., Cashel, M. Differential regulation by ppGpp versus pppGpp in Escherichia coli. Nucleic Acids Research. 41 (12), 6175-6189 (2013).
- Molodtsov, V., et al. Allosteric Effector ppGpp Potentiates the Inhibition of Transcript Initiation by DksA. Molecular Cell. 69 (5), 828-839 (2018).
- Ross, W., Sanchez-Vazquez, P., Chen, A. Y. Y., Lee, J. -. H. H., Burgos, H. L. L., Gourse, R. L. L. ppGpp Binding to a Site at the RNAP-DksA Interface Accounts for Its Dramatic Effects on Transcription Initiation during the Stringent Response. Molecular Cell. 62 (6), 811-823 (2016).
- Kriel, A., et al. Direct Regulation of GTP Homeostasis by (p)ppGpp: A Critical Component of Viability and Stress Resistance. Molecular Cell. 48 (2), 231-241 (2012).
- Kriel, A., et al. GTP dysregulation in Bacillus subtilis cells lacking (p)ppGpp results in phenotypic amino acid auxotrophy and failure to adapt to nutrient downshift and regulate biosynthesis genes. Journal of Bacteriology. 196 (1), 189-201 (2014).
- Potrykus, K., Cashel, M. p)ppGpp: still magical. Annual Review of Microbiology. 62, 35-51 (2008).
- Hauryliuk, V., Atkinson, G. C., Murakami, K. S., Tenson, T., Gerdes, K. Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nature Reviews Microbiology. 13 (5), 298-309 (2015).
- Arenz, S., et al. The stringent factor RelA adopts an open conformation on the ribosome to stimulate ppGpp synthesis. Nucleic Acids Research. 44 (13), 6471-6481 (2016).
- Loveland, A. B., et al. Ribosome•RelA structures reveal the mechanism of stringent response activation. eLife. 5, e17029 (2016).
- Brown, A., Fernández, I. S., Gordiyenko, Y., Ramakrishnan, V. Ribosome-dependent activation of stringent control. Nature. 534 (7606), 277-280 (2016).
- Battesti, A., Bouveret, E. Acyl carrier protein/SpoT interaction, the switch linking SpoT-dependent stress response to fatty acid metabolism. Molecular Microbiology. 62 (4), 1048-1063 (2006).
- Lee, J. -. W., Park, Y. -. H., Seok, Y. -. J. Rsd balances (p)ppGpp level by stimulating the hydrolase activity of SpoT during carbon source downshift in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 115 (29), E6845-E6854 (2018).
- Cashel, M. Detection of (p)ppGpp Accumulation Patterns in Escherichia coli mutants. Molecular Microbiology Techniques. 3, 341-356 (1994).
- Fernández-Coll, L., Cashel, M. Contributions of SpoT Hydrolase, SpoT Synthetase, and RelA Synthetase to Carbon Source Diauxic Growth Transitions in Escherichia coli. Frontiers in Microbiology. , (2018).
- Varik, V., Oliveira, S. R. A., Hauryliuk, V., Tenson, T. HPLC-based quantification of bacterial housekeeping nucleotides and alarmone messengers ppGpp and pppGpp. Scientific Reports. 7 (1), 11022 (2017).
- Rhee, H. -. W., et al. Selective Fluorescent Chemosensor for the Bacterial Alarmone (p)ppGpp. J. Am. Chem. Soc. 130 (3), 784-785 (2008).
- Wang, J., Chen, W., Liu, X., Wesdemiotis, C., Pang, Y. A Mononuclear Zinc Complex for Selective Detection of Diphosphate via Fluorescence ESIPT Turn-On. Journal of Materials Chemistry B. 2 (21), 3349-3354 (2014).
- Pokhilko, A. Monitoring of nutrient limitation in growing E. coli: a mathematical model of a ppGpp-based biosensor. BMC Systems Biology. 11 (1), 106 (2017).
- Funabashi, H., Mie, M., Yanagida, Y., Kobatake, E., Aizawa, M. Fluorescent monitoring of cellular physiological status depending on the accumulation of ppGpp. Biotechnology Letters. 24 (4), 269-273 (2002).
- Neidhardt, F. C., Bloch, P. L., Smith, D. F. Culture medium for enterobacteria. Journal of Bacteriology. 119 (3), 736-747 (1974).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9 (7), 671-675 (2012).
- Peselis, A., Serganov, A. ykkC riboswitches employ an add-on helix to adjust specificity for polyanionic ligands. Nature Chemical Biology. 14 (9), 887-894 (2018).
- Oki, T., Yoshimoto, A., Sato, S., Takamatsu, A. Purine nucleotide pyrophosphotransferase from Streptomyces morookaensis, capable of synthesizing pppApp and pppGpp. Biochimica et Biophysica Acta (BBA) – Enzymology. 410 (2), 262-272 (1975).
- Travers, A. A. ppApp alters transcriptional selectivity of Escherichia coli RNA polymerase. FEBS Letters. 94 (2), 345-348 (1978).
- Bruhn-Olszewska, B., et al. Structure-function comparisons of (p)ppApp vs (p)ppGpp for Escherichia coli RNA polymerase binding sites and for rrnB P1 promoter regulatory responses in vitro. Biochimica et Biophysica Acta (BBA) – Gene Regulatory Mechanisms. 1861 (8), 731-742 (2018).
