В swimmerets раков выполнять функцию в контроле позы и бить ритмично, когда животные плавать вперед, проветрите их норы или женщины проветрить свои яйца 5, 6. The swimmerets сигнала раков, Pacifastacus leniusculus, встречаются парами из второго по пятое брюшной сегмент, с одной конечности на каждой стороне брюшной полости 7. Центральной нервной системы производит на собственной ритмической двигателя стук, который приводит в движение swimmeret в интактных животных, а также в изолированном препарате нервной цепочки. Когда нет сенсорная обратная связь или убывания вход настоящее ритмичный двигатель модели производятся называется фиктивным передвижение 1, 2. В системе swimmeret этот двигатель модели не отличается какого-либо параметра от деятельности swimmerets, измеренных в интактных животных.
Движение каждого swimmeret управляется микросхемой, который находится в и ограничивается одной сorresponding hemiganglion 1 -. 3 В каждой микросхемы есть шаблон генерации ядра, включает в себя пять выявленных удобства всплески интернейронов. Они могут быть функционально характеризуется либо как ингибитор хода силы (IPS) или ингибитор обратного хода (IRS) 8. Эти IPS и IRS интернейронов не являются эндогенными генераторы, а их переменный деятельность обусловлена взаимного ингибирования 9. Потому что эти интернейронов ингибировать swimmeret моторные нейроны напрямую, переменное движение PS-RS генерируется 10. Передвижение однако, не только требуют создания деятельности, но и координации различных независимых микросхем. В системе swimmeret такая координация устанавливается координационного микросхемы, которая гарантирует, что конечности являются активными в правильное время. Эта микросхема построена на трех определенных нейронов в каждом сегменте 11-15.
Этот протокол предусматривает гое первый раз шаг за шагом рассечение руководство, чтобы изолировать цепь ганглиев (T4 до A6, рис 1). Мы покажем, как прикрепить Изолированная абдоминальная нервный тяж и desheathe каждый ганглий. В этом изолированном препарате нервной системы, нейроны, отвечающие за движения swimmeret готовы для использования в электрофизиологических и морфологических экспериментов. Вторая часть этого протокола демонстрирует основные особенности swimmeret узором двигателя. Это включает в себя шаг за шагом руководство для внеклеточно записи активности swimmeret двигательных нейронов. Аксоны двигательных RS нейроны проецируются через передней ветви нерва N1, в то время как аксоны PS двигательных нейронов проекта через заднюю ветви одного и того же нерва (рис 1) 4. Поэтому их деятельность может быть записано от этих отраслей с помощью дифференциальных контактных электродов.
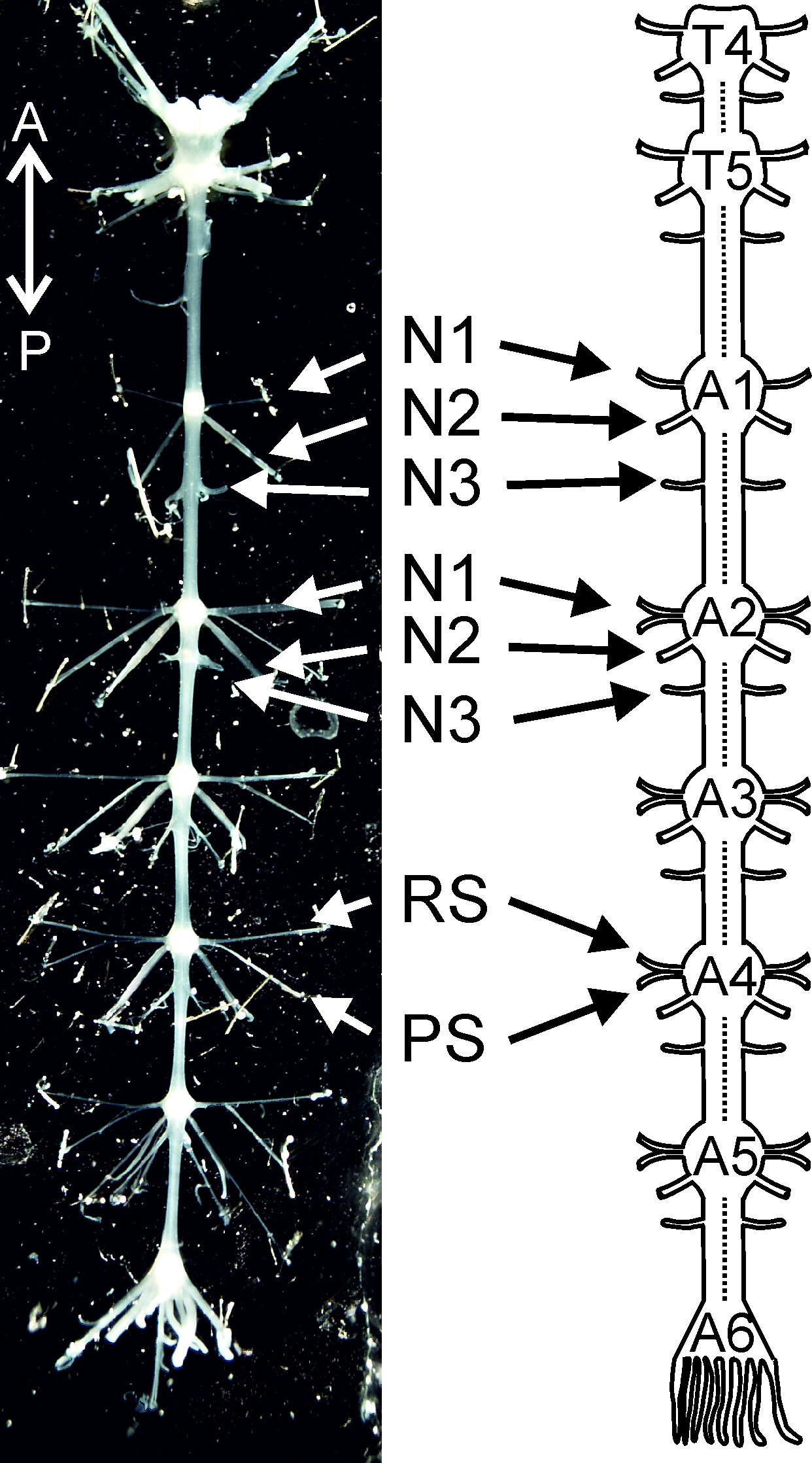 <br/> Рисунок 1: Изолированные нервной системы от грудного ганглия 4 (Т4) в брюшной ганглий 6 (A6) и принципиальная схема, что Т4.: Грудной ганглий 4; T5: грудной ганглий 5; A1, A2 … A6 брюшной ганглий 1, брюшной ганглий 2 … брюшной ганглий 6; N1: нерв N1; N2: нерв N2; N3: нерв N3; PS: рабочий ход; RS: возвращение-тактный. Направленные сокращения: = Передняя; P = задний.
<br/> Рисунок 1: Изолированные нервной системы от грудного ганглия 4 (Т4) в брюшной ганглий 6 (A6) и принципиальная схема, что Т4.: Грудной ганглий 4; T5: грудной ганглий 5; A1, A2 … A6 брюшной ганглий 1, брюшной ганглий 2 … брюшной ганглий 6; N1: нерв N1; N2: нерв N2; N3: нерв N3; PS: рабочий ход; RS: возвращение-тактный. Направленные сокращения: = Передняя; P = задний.
Эта процедура рассечение и электрофизиологические техника продемонстрировала удобны для студентов и могут дополнять студентов практические занятия по физиологии. Изолированные цепи ганглиев был использован в ряде экспериментов по изучению функции нервной системы, координацию, или модуляцию swimmeret микросхем 6, а также нейронную контроль адаптивного поведения в локомоции 16, 17. Таким образом, система раков swimmeret обеспечивает огромное количество интересной преподавательской или тдождь возможности, которые все начинаются с рассечением брюшной нервной раков и внеклеточной записи фиктивного узором двигателя.
