Adenosine (ADO) est une purine nucléoside avec une molécule d'adénine attachée à un fragment de molécule de sucre ribose par une liaison glycosidique. Lorsqu'ils sont présents dans le milieu extracellulaire, elle protège les cellules contre les dommages excessifs par l'action du système immunitaire. Ce rôle a été mis en évidence en utilisant différents modèles de maladies, telles que la colite 1, 2 diabète, l' asthme 3, la septicémie 4 et 5 lésion ischémique. L' une des principales fonctions d'ADO est l'inhibition des réponses immunitaires dans le microenvironnement de la tumeur, ce qui contribue à la tumeur évasion immunitaire 6. Pour cette raison, les mécanismes impliqués dans la formation ADO et la signalisation sont d' un intérêt thérapeutique considérable 7.
ADO niveaux dans le microenvironnement des tissus sont relativement faibles dans des conditions physiologiques normales, et certainement inférieure au seuil de sensibilité des cellules immunitaires. Cependant, au cours de l'hypoxie, l'ischémie, l'inflammation, l'infection, métaboliquele stress et la transformation tumorale , ils augmentent rapidement 8. Les niveaux d'ADO extracellulaires élevées en réponse à des signaux de tissu perturbant ont une double fonction: signaler une lésion tissulaire de façon autocrine et paracrine et pour générer des réponses tissulaires qui peuvent être généralement considérés comme cytoprotecteur.
Extracellulaires ADO peut être formé par une variété de mécanismes, qui comprennent la libération des compartiments intracellulaires médiés par les transporteurs de nucléosides 9 ou accumulation en raison de la dégradation altérée exploité par l' adénosine désaminase. La voie principale menant à une augmentation des niveaux extracellulaires ADO implique l'action d'une cascade de ectonucléotidases, qui sont associées à la membrane ectoenzymes générant ADO par phosphohydrolysis de nucléotides libérés de cellules mortes ou mourantes. Cette voie passe par l'action séquentielle de CD39 (triphosphate ectonucleoside diphosphohydrolase-1) qui convertit l'adénosine extracellulaire 5'-triphosphate (ATP) ou l' adénosine 5'-diphosphate (ADP) en adénosine 5'-monophosphate (AMP) et de CD73 (5'-nucléotidase) qui transforme l' AMP à 10 ADO.
Extracellulaires ADO provoque ses réactions physiologiques en se liant à quatre transmembranaire ADO récepteurs, à savoir A1, A2A, A2B et A3. Chaque récepteur a des affinités différentes pour ADO et la distribution des tissus spécifiques. Tous les récepteurs ont sept domaines transmembranaires et sont couplés aux protéines G à des protéines de liaison au GTP intracellulaire (protéines G), qui peut induire (protéine Gs) ou à inhiber (Gi protéine) activité de l'adénylate cyclase et, par la suite, la production d'AMPc intracellulaire. Par conséquent, les changements de l' impact cytoplasmique des taux d'AMPc sur l' activité protéine kinase intracellulaire pendant 11 réponses physiologiques. Dans des conditions physiologiques ADO extracellulaire est inférieure à 1 uM, qui peut activer indifféremment A1, A2A et récepteurs A3. Cependant, l'activation du sous-type A2B nécessite considérablement plus élevésLes concentrations du nucleoside, tels que ceux produits dans des conditions physiopathologiques. En variante, OAD extracellulaire peut être dégradée à l'inosine (INO) par l'adénosine désaminase (ADA) et de CD26, une protéine ADA complexant localisant l'ADA sur la surface cellulaire. Une autre possibilité est que ADO est internalisé par la cellule par les transporteurs de nucléosides (ORL) et phosphorylée à AMP par ADO protéine kinase 12,13.
L'objectif de ce protocole est de décrire une méthode d'analyse de la chromatographie en phase liquide à haute performance en phase inverse (RP-HPLC) pour quantifier en un seul passage AMP du substrat et des produits ADO et INO, telle que générée par les lymphocytes humains. Notre expérience a été initialement obtenue en utilisant des cellules de patients chroniques leucémie lymphoïde (CLL), qui sont caractérisés par l'expansion d'une population mature de lymphocytes CD19 + / CD5 + B exprimant constitutivement CD39 14,15. Nous avons montré environ 30%des CLL patients expriment le ectoenzyme CD73 et que ce phénotype est corrélée à un mauvais pronostic 16. Cette sous-population de cellules leucémiques co-exprimant CD39 et CD73 peut activement produire ADO extracellulaire d'ADP et / ou AMP. La pré – incubation de cellules CD73 + leucémiques avec des α, la synthèse d'ADO extracellulaire β-méthylène-ADP (PCAP), un inhibiteur connu de l' activité enzymatique CD73, bloque complètement confirmant que l'enzyme CD73 représente un goulot d'étranglement de cette cascade 16.
les cellules CLL expriment également ADA et l'ADA protéine complexant CD26, qui sont responsables de la conversion de ADO dans INO. En utilisant des inhibiteurs de l'ADA spécifiques, tels que érythro-9- (2-hydroxy-3-nonyl) Je wiadenine (EHNA) et du chlorhydrate désoxycoformycine (FNC), il est possible de bloquer la dégradation ADO extracellulaire dans INO. En outre, le prétraitement avec un inhibiteur de l'ADA, en combinaison avec le dipyridamole, qui bloque les transporteurs de nucléosides, augmente l'accumulation dans la cellule ADOsurnageants.
Nous avons ensuite étendu ce protocole à des cellules dérivées d'autres lignées, y compris les lymphocytes T et les cellules myéloïdes, confirmant la production d'ADO CD73-dépendante. Ces résultats suggèrent que ce protocole HPLC est très polyvalent et qu'il peut être appliqué à différentes lignées cellulaires et à différentes conditions de culture (figure 1).
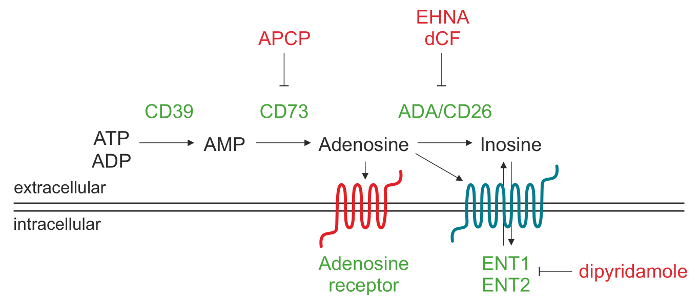
Figure 1. Représentation schématique de la machinerie enzymatique responsable de la production ADO extracellulaire. Adenosine 5'-triphosphate (ATP) et / ou l' adénosine 5'-diphosphate (ADP) peut être dégradé par CD39 à l' adénosine 5'-monophosphate (AMP), qui à son tour, est converti par CD73 dans l'adénosine nucléoside (ADO). Une fois que ADO est produite dans l'espace extracellulaire, elle peut rentrer dans la cellule par les transporteurs de nucléosides (ENT), être converti en l'inosine (INO) oulier à différents types de récepteurs P1 ADO. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
