自 1873年, 由于来自目标12的光线衍射, 传统光学显微镜的分辨率被认为仅限于大约 200 nm。目前, 超分辨率光显微镜技术打破了这一限制, 并允许捕捉动态图像与 sub-diffraction (< 200 nm) 的决议。该技术一般分为两大类: 受激发射损耗 (STED) 显微镜的方法, 它产生 sub-diffraction 光照体积由于非线性光学响应的荧光在样本3;和光光显微镜 (手掌) 和随机光学重建显微镜 (风暴) 的超分辨率技术, 利用数学功能, 本地化质心的荧光, 然后重建这些质心要形成超分辨率图像4,5。目前, 由于相对简单的光学设置, 棕榈和风暴是广泛使用的只激活一个小子集的荧光在每个帧的一个长视频的生物准备。这使得更精确的定位由2D 高斯拟合的荧光点, 称为点扩散函数 (聚砜), 荧光标记的蛋白质在每个帧的视频。每个荧光标记的分子的2D 位置, 然后可以叠加在一个单一的成像平面上产生一个超分辨率的图像的生物准备1,2。虽然这些分子的本地化, 超分辨率的方法, 以显微镜肯定革命性的生物样品的成像是如何进行的, 仍然有挑战要克服。例如, 风暴和棕榈可以达到他们的最佳空间分辨率后, 固定的生物样品, 从而提出了静态表示的荧光标记蛋白, 这是一个类似的限制电子显微镜。此外, 为了实现在活细胞中每个荧光标记的蛋白质的高空间分辨率, 样品必须成像在非常长的帧, 不能捕捉蛋白质动力学。因此, 有必要克服这些主要的技术障碍。
为了获得一个高时空分辨率, 它非常适合在活细胞中检测快速移动的蛋白质或 rna, 我们在实验室中开发了超分辨速度显微镜 (图 1)6,7,8. 速度显微镜的几个主要技术进步使我们能够成功地追踪小分子、蛋白质、mRNA 和病毒通过本机核孔复合体 (npc) 的 nucleocytoplasmic 传输,6,7,8. 简要地, 以下速度显微镜的特点将用于跟踪快速移动的大分子通过微米旋转对称结构在活细胞, 例如 npc 和主要纤毛: (1) 倾斜或垂直照明, 使在焦平面的小衍射极限体积内的单分子激发 (图 1);(2) 倾斜的聚砜可以极大地避免焦荧光, 从而提高信噪比。(3) 光照中的 100-500 kW/cm2的光学密度允许数千个光子从单个荧光中收集, 并具有快速检测速度 (> 500 Hz)。(4) 快速检测速度还大大降低了分子空间定位错误 (< 10 nm) 在确定活细胞中移动荧光分子的空间轨迹时, 因为分子扩散是主要因素之一造成移动分子分子定位的缺陷。(5) 良好的2D 到3D 转换算法使我们能够为 NPC 或主要纤毛中的分子提供3D 空间概率密度地图。值得注意的是, 我们在笛卡尔和圆柱坐标系之间的转换过程被用来生成3D 空间概率密度图而不是3D 分子跟踪 (图 2)。此前, 电子显微镜数据显示, NPC9、10和主纤毛11都具有旋转对称结构。原则上, 随机扩散分子通过 NPC 或初级纤毛也应该有旋转对称分布。如图 2所示, 大量随机扩散的分子在气缸内将产生旋转对称分布, 在剖面图, 在 NPC, 进一步导致一个近似一致的空间在相邻的两个环之间的每个非常小的子区域中分布 (图 2E)。这种均匀分布导致圆柱系统中θ维数的空间分布是恒定的。然后, 3D 坐标 (r、x、θ) 可以简化为2D 坐标 (r、x、常量)。实际上, 我们在笛卡尔和圆柱系统之间的转换过程是从 2D (x, Y) 到 2D (R, x, 常数)。常数θ, 指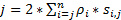 的是空间密度p在图 2E中, 是通过使用等式A来计算的。
的是空间密度p在图 2E中, 是通过使用等式A来计算的。
最终, 分子跟踪在生物研究中有着广泛的应用, 因此, 将开发大量的技术来填充特定的生物龛位12、13、14是很自然的。速度显微镜就是如此。以前, 当与3D 变换算法结合时, 这项技术被开发来解决通过 npc 传递分子的3D 传输路线, 一个 sub-diffraction-sized 和旋转对称的生物结构6。本文中, 原发纤毛也被证明是优秀的模型细胞器。主要纤毛是圆柱形的, 天线状的细胞器 (~ 125 nm 半径), 该项目从大多数哺乳动物的表面15,16,17。它们负责接收外部信号, 并传送通常与生长和新陈代谢相关的细胞内反应15,16。因此, 结构蛋白的通量、跨膜受体的再循环以及细胞内信使的传递是原发性纤毛的重要责任。在主要纤毛和细胞体之间的接合处是一种临界选择性屏障, 称为过渡区或正向, 所有这些蛋白传输必须发生11,18,19, 20。除了 intraflagellar 的浇注功能, 至少有两个传输过程, 即传输和被动扩散, 被认为是负责蛋白质的移动通过这个区域16,21,22. 从人类健康的角度来看, 失去初级纤毛和随后对下游信号的放松管制是许多癌症的特点。此外, 许多遗传性疾病, 如 Bardet Biedl 综合征和多囊肾病, 都与有缺陷的蛋白质转运有关23。sub-diffraction 极限大小和选择性蛋白转运的复杂过程, 使原纤毛成为这项技术的主要目标。在本方法中, 我们将展示一个睫状跨膜蛋白的跟踪, 生长抑素受体 3 (SSTR3)24, 在外部标有 Alexa 氟647和 IFT, IFT2025的成分, 用一个融合的 GFP 分子标记。
