Так как заявил Эрнст Аббе в 1873 году, резолюции обычных световой микроскопии было считается ограничивается примерно 200 Нм вследствие дифракции света от объективного1,2. В настоящее время методы световой микроскопии суперразрешением нарушать это ограничение и разрешить захват динамических изображений с разрешением суб дифракции (< 200 Нм). Методы, как правило, делятся на две широкие категории: стимулировали выбросов истощения (интереса) микроскопии основанные подходы, которые генерируют объем освещения суб дифракции вследствие нелинейных оптических ответ флуорофоров в образцы3; и photoactivated световой микроскопии (PALM) и стохастические оптических реконструкции микроскопии (шторм)-на основе суперразрешением методы, которые используют математические функции для локализации центроиды флуорофоров и затем восстановить эти центроиды сформировать суперразрешением изображения4,5. В настоящее время благодаря сравнительно несложной оптические установки, PALM шторм широко используются и активируя только небольшое подмножество флуорофоров в каждом фрейме долго видео биологической подготовки. Это позволяет для более точной локализации по 2D Гаусса фитинга флуоресцентные пятна, называется точка распространения функция (ФСФ), дневно меченых белков в каждом кадре видео. 2D расположение каждого дневно меченых молекул может затем накладывается на одной плоскости изображений для создания изображения суперразрешением биологической подготовки1,2. Хотя эти сингл молекула локализации, супер резолюции подходы к микроскопии конечно революцию, как было выполнено визуализации биологических образцов, есть еще проблемы, которые необходимо преодолеть. Например шторм и PALM могут достигнуть их лучших пространственного разрешения после фиксации биологических образцов и таким образом представлять статическое представление дневно меченых белков, который является аналогичные ограничения электронной микроскопии. Кроме того для достижения высокого пространственного разрешения для каждого белка-дневно обозначенных в живых клеток, образцы должны быть imaged на очень длинные частоты кадров, которые не в состоянии захватить динамики белков. Таким образом это необходимо для преодоления этих основных технических препятствий.
Чтобы получить высокое разрешение пространственно-временных, которая хорошо подходит для обнаружения быстро движущихся белков и РНК в живых клеток, мы разработали супер-резолюции скорость микроскопии в нашей лаборатории (рис. 1)6,7, 8. несколько крупных технических достижений в микроскопии скорость ранее позволили нам успешно отслеживать nucleocytoplasmic перевозок малых молекул, белки, мРНК и вирус через родной ядерной поры комплексы (НПС)6, 7 , 8. кратко, следующие особенности микроскопии скорость будет использоваться для отслеживания быстро движущихся макромолекул через суб микрометра осесимметричной структуры в живых клеток, таких как NPC и первичных реснички: (1) наклонной или вертикальная подсветка ПСФ позволяет возбуждения молекул одного внутри небольшой дифракционный предел тома в фокальной плоскости (рис. 1); (2) склонны ПСФ можно значительно избежать вне фокуса флуоресценции и таким образом улучшить соотношение сигнал шум. (3) оптическая плотность 100-500 кВт /2 см в ПСФ освещения позволяет тысячи фотоны должны быть собраны из одного флуорофоров с быстрого обнаружения скорости (> 500 Гц). (4) быстрого обнаружения скорость также значительно уменьшает сингл молекула пространственной локализации ошибки (< 10 Нм) при определении пространственных траекторий перемещения флуоресцентных молекул в живых клеток, потому что молекулярной диффузии является одним из основных факторов причиной несовершенства одной молекулы локализации для движущихся молекул. (5) налаженные 2D к 3D алгоритмы преобразования позволяют нам предоставлять 3D пространственных вероятностных карт плотности транспортных маршрутов для молекул в NPC или первичной ресничку. Примечательно, что наш процесс преобразования между декартовой и цилиндрических координации системы используется для создания 3D пространственной плотности вероятности карта, вместо того, чтобы 3D сингл молекула отслеживания (рис. 2). Ранее электронной микроскопии данные показали, что ВСНП9,10 и основной ресничку11 имеют осесимметричной структуры. В принципе случайно диффундирующих молекул, движущихся через NPC или первичной ресничку должны также иметь осесимметричной дистрибутивов. Как показано на рисунке 2, большое количество случайно диффундирующих молекул внутри цилиндра будет генерировать осесимметричной дистрибутивов на вид поперечного сечения, что в NPC, далее привело примерно единообразной пространственной распределение в рамках каждого очень маленькие субрегиона между двумя соседними кольцами (2 рисунокE). Это равномерное распределение приводит, что пространственное распределение измерении θ в цилиндрической системе является постоянным. Затем можно упростить 3D координаты (X, R, θ) быть 2D координат (R, X, константа). На самом деле наш процесс преобразования между декартовой и цилиндрических системами является от 2D (X, Y) 2D (R, X, константа). Постоянное θ, относится к пространственной плотности p в рисунке 2E, рассчитывается с помощью уравнения A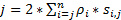 .
.
В конечном счете сингл молекула отслеживания имеет широкое применение в биологических исследований, таким образом, вполне естественно, что множество методов будут разрабатываться для заполнения конкретных биологических нишах12,,1314. Так обстоит дело с скорость микроскопии. Ранее при сочетании с 3D преобразования алгоритма, этот метод был разработан для устранения 3D транспортных маршрутов транзитных молекул через НПК, суб-diffraction-размера и вращательно-симметричных биологической структуры6. В этом документе первичный ресничек показано отличную модель органеллы также. Основная ресничек, цилиндрические, антенна как органеллы (~ 125 Нм радиус), которые проектируют от поверхности наиболее mammalian клетки15,16,17. Они отвечают за получение внешних сигналов и препровождающее внутриклеточные реакции, обычно связанных с ростом и метаболизм,1516. Таким образом, поток структурных белков, утилизация трансмембранные рецепторы и передаче внутриклеточных посланников жизненно важные обязанности основного ресничек. На стыке между первичной ресничек и клетки тела является критической избирательности барьер, называется переходной зоне или TZ, через который все это транспортного белка должно произойти,11,18,19 20. Подуманы, что помимо стробирования функции TZ, по крайней мере два транспортных процессов, intraflagellar транспорт и пассивной диффузии, отвечает за движение белка через этот регион16,21, 22. с точки зрения здоровья человека, потеря первичных ресничек и последующего дерегулирования течению сигнализации характерно многих раковых заболеваний. Кроме того многие генетических заболеваний, как Синдром Барде-Бидля и поликистозный болезни почек, связаны с дефектных белков транспорт23. Ограничение на размер суб дифракции и сложный процесс селективного белка транспорта через TZ сделать основной реснички главной мишенью для этой техники. В этом документе методы, мы продемонстрируем отслеживания цилиарной трансмембранным белком рецепторов соматостатина 3 (SSTR3)24, внешне помечены Alexa Fluor 647 и компонент IFT, IFT2025, помечены с молекулой плавленого GFP.
