מאז כאמור על ידי ארנסט אבה בשנת 1873, הרזולוציה של מיקרוסקופ אור קונבנציונלי יש כבר האמין להיות מוגבל כ 200 ננומטר עקב שבירת האור אובייקטיבית1,2. כיום, סופר-רזולוציה מיקרוסקופ אור טכניקות לשבור מגבלה זו ולאפשר לכידת תמונות דינאמית ברזולוציה תת עקיפה (< 200 ננומטר). הטכניקות בדרך כלל מתחלקים לשתי קטגוריות רחבות: גירוי פליטה דלדול (STED) מיקרוסקופיה המבוסס על גישות, המניבות עקיפה תת נפח תאורה עקב תגובת אופטית לא-לינאריות fluorophores דגימות3; מיקרוסקופ אור photoactivated (דקל), מיקרוסקופיה סטוכסטי שחזור אופטי (סערה)-המבוסס על שיטות זיהוי סופר, אשר מנצלים את פונקציות מתמטיות, ולהתאימם לעגה של centroids של fluorophores ואז לשקם את centroids האלה כדי ליצור סופר רזולוציה תמונות4,5. כיום, עקב הגדרת אופטי מסובכת יחסית, סערה ודקלים בהרחבה מועסקים על ידי הפעלת רק קבוצת משנה קטנה של fluorophores בכל מסגרת של סרטון ארוך של תכשיר ביולוגי. זה מאפשר עבור ההתאמה מדויקת יותר על ידי התאמה לפי עקומת גאוס 2D של הנקודה פלורסנט, כינה את הפונקציה הליין (PSF), שכותרתו fluorescently חלבונים בכל מסגרת של הוידאו. המיקום 2D של כל מולקולה עם התווית fluorescently ואז יכול להיות יונחו על מטוס הדמיה בודד כדי לייצר תמונת סופר רזולוציה של הכנה ביולוגי1,2. בזמן לוקליזציה מולקולה בודדת אלה, גישות סופר-רזולוציה מיקרוסקופיה בהחלט מהפכה כיצד בוצעה הדמיה של דגימות ביולוגיות, עדיין ישנם אתגרים להתגבר. לדוגמה, סערה ודקל ניתן להשיג החלטות המרחבי בצורה הטובה ביותר שלהם לאחר קיבוע של דגימות ביולוגיות, ובכך להציג ייצוג סטטי של החלבונים עם התווית fluorescently, אשר היא מגבלה דומה של מיקרוסקופ אלקטרונים. בנוסף, להשיג רזולוציה מרחבית גבוהה עבור כל חלבון הנקרא fluorescently בתאים חיים, דוגמאות חייב לדימות-framerates רב אשר אינם מסוגלים ללכוד dynamics חלבון. לכן, יש צורך להתגבר על מכשולים טכניים הראשי אלה.
כדי להשיג רזולוציה גבוהה ייתכן כי הוא מתאים היטב עבור זיהוי מהיר-המעבר חלבונים או RNAs בתאים חיים, פיתחנו רזולוציה סופר מהירות מיקרוסקופ ב שלנו מעבדה (איור 1)6,7, 8. ההתקדמות הטכנית מרכזיים במיקרוסקופ מהירות בעבר אפשרו לנו לעקוב בהצלחה nucleocytoplasmic התחבורה של מולקולות קטנות, חלבונים, mRNA ווירוסים דרך מקורית גרעיני הנקבוביות מתחמי (NPCs)6, 7 , 8. בקצרה, התכונות הבאות של מהירות מיקרוסקופ ישמש למעקב מהיר-המעבר מקרומולקולות דרך מבנים סימטריים rotationally מיקרומטר תת בתאים חיים, כגון NPCs cilia ראשי: (1) נוטה או תאורה אנכי PSF מאפשר את עירור של מולקולות יחיד בתוך אמצעי אחסון קטן עקיפה הגבלה ב מישור מוקד (איור 1); (2) PSF נוטה ניתן במידה רבה למנוע out-of-להתמקד פלורסצנטיות, ובכך לשפר את יחס אות לרעש. (3) הצפיפות האופטית של 100-500 קילוואט / ס מ2 ההארה PSF מאפשר אלפי פוטונים להיות שנאספו מ- fluorophores אחד עם זיהוי מהר למהירויות (> 500 Hz). (4) המהיר זיהוי מהירות מקטינה באופן משמעותי גם את השגיאה לוקליזציה המרחבי מולקולה בודדת (< 10 ננומטר) בקביעת את מסלולי המרחבי של מולקולות פלורסנט לגור תאים חיים, כי פעפוע הוא אחד הגורמים העיקריים גורם פגמים של מולקולה בודדת לוקליזציה להעברת מולקולות. (5) להקציע 2D ל 3D המרה אלגוריתמים מאפשרים לנו לספק מפות צפיפות ההסתברות מרחבית תלת-ממד של נתיבי תחבורה עבור מולקולות של NPC או את ריסי העיקרי. ראוי לציין כי תהליך ההמרה בין את קרטזי לבין מערכת תיאום גלילי משמש ליצירת צפיפות ההסתברות מרחבית תלת-ממד מפה ולא תלת-ממד יחיד מולקולה מעקב (איור 2). בעבר, מיקרוסקופ אלקטרונים נתונים נחשפו9,NPC10 ו ריסי הראשי11 לשניהם יש מבנה סימטרי rotationally. בעקרון, באופן אקראי לשדר המולקולות דרך NPC או ריסי הראשי צריך גם הפצות rotationally סימטרי. כפי שמוצג באיור2, מספר גבוה של אקראי לשדר מולקולות בתוך הגליל ליצור הפצות סימטרית rotationally-תצוגת חתך הרוחב עד כדי כך ב NPC, נוסף והתוצאה אחידה כ מרחבית הפצה בכל אזור משנה קטן מאוד בין שתי השכנות טבעות (איור 2E). התפלגות אחידה זה מוביל התפוצה המרחבית לאורך ממד θ באמצעות במערכת צירים היא קבועה. לאחר מכן ניתן לפשט את קואורדינטות תלת-ממד (R, X, θ באמצעות) להיות הקואורדינטות 2D (R, X, קבוע). למעשה, תהליך ההמרה בין קרטזי את המערכות גלילי הוא מ- 2D (X, Y) 2D (R, X, קבוע). Θ באמצעות מתמדת, מתייחס הצפיפות המרחבי p ב- איור 2E, מחושב באמצעות המשוואה A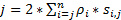 .
.
בסופו של דבר, מולקולה בודדת המעקב מצא יישום רחבות במחקר ביולוגי, לכן טבעי כי שפע של טכניקות יתפתחו למלא נישות ביולוגי מסוים12,13,14. כזה הוא המקרה עם מיקרוסקופ מהירות. בעבר, כאשר בשילוב עם אלגוריתם שינוי תלת-ממד, טכניקה זו פותחה כדי לפתור נתיבי תחבורה תלת-ממדי של מולקולות דרך NPCs, משנה-diffraction בגודל rotationally סימטרי ביולוגי ומבנה6הם מגיעים. בנייר זה, מוצגים cilia הראשי להיות דגם מעולה organelles גם כן. Cilia הראשי הם גלילי, כמו אנטנה organelles (~ 125 nm radius) המקרינים מפני השטח של ביותר בתרבית של תאים15,16,17. הם אחראים על קבלת אותות חיצוניים והעברת מענה תאיים הקשורים בדרך כלל צמיחה חילוף החומרים15,16. לכן, שטף של חלבונים מבניים, מיחזור של רצפטורים transmembrane, והעברת שליחי תאיים האחריות חיוני של cilia העיקרי. בצומת בין של cilia העיקרי לגוף התא היא מכשול סלקטיביות קריטיים, המכונה אזור המעבר או צ’, שדרכו כל העברה חלבון זו חייבת להתבצע11,18,19, 20. בנוסף הפונקציה חסימה של למתחם, נחשבים לפחות שני תהליכים התחבורה, תחבורה intraflagellar, דיפוזיה פסיבית, להיות אחראי על התנועה של חלבון דרך זה אזור16,21, 22. מבחינת בריאות האדם, האובדן של cilia העיקרי, ורשות עוקבות של איתות במורד הזרם היא תכונה של סרטן רבים. בנוסף, מחלות גנטיות רבות, כגון תסמונת Bardet-Biedl, מחלת כליות פוליציסטיות, משויכים חלבון פגום תחבורה23. עקיפה תת מגבלת גודל והן את תהליך מורכב של חלבונים סלקטיבי התחבורה באמצעות למתחם להפוך cilia העיקרי זה ליעד עיקרי עבור טכניקה זו. בנייר זה שיטות, נדגים את המעקב אחר טראנסממברנלי ciliary, קולטני סומטוסטטין 3 (SSTR3)24, תווית חיצונית עם אלקסה עבור חיל הים 647, רכיב של: אי, IFT2025, המסומנת מולקולה GFP מאוחה.
