Очистка и контроль качества рекомбинантных комплексов септина для бесклеточного восстановления
Summary
Восстановление in vitro цитоскелетных белков является жизненно важным инструментом для понимания основных функциональных свойств этих белков. В настоящем документе описывается протокол очистки и оценки качества рекомбинантных комплексов септина, которые играют центральную роль в делении и миграции клеток.
Abstract
Септины представляют собой семейство законсервированных эукариотических GTP-связывающих белков, которые могут образовывать цитоскелетные нити и структуры более высокого порядка из гетероолигомерных комплексов. Они взаимодействуют с другими компонентами цитоскелета и клеточной мембраной для участия в важных клеточных функциях, таких как миграция и деление клеток. Из-за сложности многих взаимодействий септинов, большого количества генов септина (13 у людей) и способности септинов образовывать гетероолигомерные комплексы с различным составом субъединиц бесклеточное восстановление является жизненно важной стратегией для понимания основ биологии септина. В настоящей работе впервые описан способ очистки рекомбинантных септинов в их гетероолигомерной форме с использованием двухступенчатого подхода аффинной хроматографии. Затем подробно описывается процесс контроля качества, используемый для проверки чистоты и целостности комплексов септина. Этот процесс сочетает в себе нативный и денатурирующий гель-электрофорез, электронную микроскопию с отрицательным пятном и интерферометрическую микроскопию рассеяния. Наконец, дано описание процесса проверки полимеризационной способности септиновых комплексов с помощью электронной микроскопии отрицательных пятен и флуоресцентной микроскопии. Это свидетельствует о том, что можно производить высококачественные гексамеры и октамеры септина человека, содержащие различные изоформы septin_9, а также гексамеры Drosophila septin.
Introduction
Цитоскелет был классически описан как трехкомпонентная система, состоящая из актиновых нитей, микротрубочек и промежуточных нитей1, но недавно септины были признаны четвертым компонентом цитоскелета1. Септины представляют собой семейство GTP-связывающих белков, которые сохраняются в эукариотах2. Септины участвуют во многих клеточных функциях, таких как деление клеток3, клеточная адгезия4, подвижность клеток5, морфогенез6, клеточная инфекция7, а также установление и поддержание полярности клеток8. Несмотря на их важные функции, как септины участвуют в таких процессах, плохо изучено.
Семейство белков септина подразделяется на несколько подгрупп (четыре или семь, в зависимости от классификации) на основе сходства последовательностей белков2. Члены различных подсемейств могут образовывать палиндромные гетероолигомерные комплексы, которые являются строительными блоками нитей и которые, в свою очередь, собираются в структуры более высокого порядка, такие как пучки, кольца и сетки 1,9,10,11,12. Дальнейшая молекулярная сложность возникает из-за наличия различных вариантов сращивания, примером чего является человеческий SEPT9, где есть доказательства специфических функций различных вариантов сращивания 13,14,15. Кроме того, длина гетероолигомеров зависит от вида и типа клеток. Например, септины Caenorhabditis elegans образуют тетрамеры16, септины Drosophila melanogaster образуют гексамеры17 (рисунок 1A), saccharomyces cerevisiae septins образуют октамеры18, а человеческие септины образуют как гексамеры, так и октамеры19 (рисунок 1A). Способность изоформ септина, вариантов сращивания и посттрансляционно модифицированных септинов из одного и того же подсемейства заменять друг друга в комплексе и (со)существование гетероолигомеров разного размера затрудняют разграничение клеточных функций различных гетероолигомерных комплексов12.
Еще одной интересной способностью септинов является их способность взаимодействовать со многими партнерами по связыванию в клетке. Септины связывают плазматическую мембрану и мембранные органеллы во время интерфазы и деления клеток 20,21,22. В делящихся клетках септины взаимодействуют с аниллином 23,24,25 и актином и мионином во время цитокинеза 26,27. На поздних стадиях цитокинеза септины, по-видимому, регулируют эндосомальные сортировочные комплексы, необходимые для транспортной системы (ESCRT) для абсциссии среднего тела28. Кроме того, имеются также данные о септине, расположенном на актиновой коре и актиновых стрессовых волокнах клеток в межфазных клетках 29,30,31. В определенных типах клеток септины также связывают и регулируют микротрубочки цитоскелета32,33.
Все эти особенности делают септины очень интересной белковой системой для изучения, но также и сложной. Сочетание большого количества субъединиц септина (13 генов у человека без учета вариантов сращивания2) с потенциалом субъединиц септина из одного и того же подсемейства заменять друг друга и образовывать гетероолигомеры разного размера затрудняет вывод о клеточной функции конкретного септина путем генетических манипуляций. Кроме того, множественные взаимодействия септинов делают интерпретацию эффектов общих исследовательских инструментов, таких как препараты34 , направленные на цитоскелетные или мембранные компоненты, трудной задачей.
Способ преодолеть эту ситуацию заключается в том, чтобы дополнить исследования в клетках восстановлением септинов in vitro (бесклеточным). Восстановление in vitro позволяет выделить один тип гетероолигомеров септина с определенным субъединицей и длиной 18,35,36,37. Затем этот комплекс может быть изучен в контролируемой среде, либо в одиночку, чтобы обнаружить основные структурные и физико-химические свойства септинов 38,39,40, либо в сочетании с желаемыми партнерами, такими как модельные биомембраны 11,41,42, актиновые нити 10,27 или микротрубочки 32,36, чтобы расшифровать природу их Взаимодействия.
Поэтому надежный метод эффективной очистки различных комплексов септина жизненно важен для исследования септина. Однако, даже используя один и тот же протокол, разные очистки могут давать белки с разной активностью / функциональностью или даже целостностью. Для коммерчески доступных белков, таких как ферменты, функциональность и ферментативная активность тщательно проверяются43. Осуществление тщательного контроля качества цитоскелетных или структурных белков, таких как септины, может быть сложной задачей, но важно сделать эксперименты в лабораториях сопоставимыми.
В данной работе описан надежный метод очистки высококачественных рекомбинантных септинов в их гетероолигомерной форме, основанный на одновременной экспрессии двух векторов, содержащих моно- или бицистронные конструкции (таблица 1) в клетках кишечной палочки. Метод состоит из двухэтапного подхода аффинной хроматографии для захвата гетероолигомеров септина, содержащих как его 6-меченый септин, так и септин со стрептиком-II (рисунок 1B, C). Этот протокол, впервые описанный в Iv et al.10, был использован для очистки гексамеров септина Drosophila 11,27,35, гексамеров септина человека 10 и нескольких октамеров септина человека, содержащих различные нативные (изоформы 1, 3 и 5)10,32 или мутировавшие изоформы SEPT932 . Кроме того, дается описание набора методов оценки качества очищенных септинов. Во-первых, целостность и правильная стехиометрия субъединиц септина проверяется с помощью денатурирующего электрофореза и просвечивающей электронной микроскопии (ТЭМ). Затем наличие гетероолигомеров правильной молекулярной массы и наличие мономеров или более мелких олигомеров, указывающих на сложную нестабильность, исследуют с помощью нативного электрофореза и массовой фотометрии с помощью интерферометрической рассеянной микроскопии (iSCAT). Наконец, последний этап состоит из оценки полимеризующей активности септинов с помощью флуоресцентной микроскопии и ТЭМ.
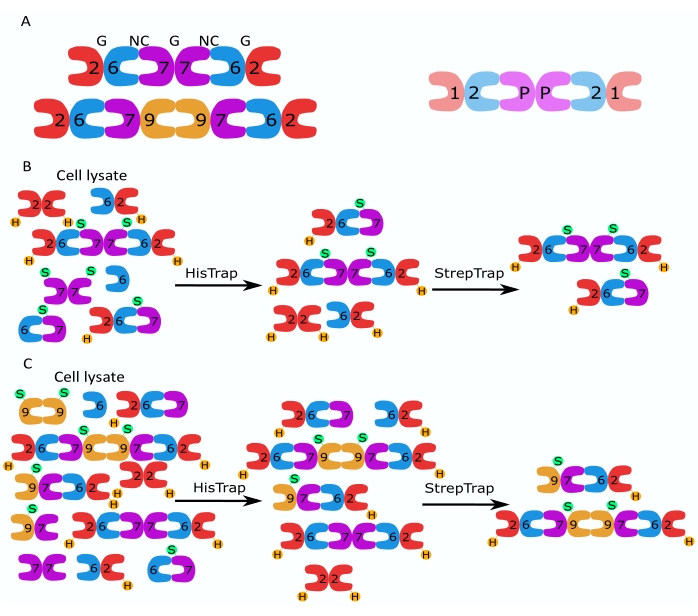
Рисунок 1: Стратегия очистки. (А) Схемы гетероолигомеров септина, которые существуют в клетках человека (слева) и дрозофилы (справа). Числа обозначают субъединицы септина из указанных групп, а P обозначает арахис. Человеческий SEPT9 может быть любой из его изоформ. Субъединицы септина имеют асимметричную форму и продольно связаны с двумя различными интерфейсами, NC:NC и интерфейсом G:G, обозначаемыми NC и G, соответственно, поверх человеческого гексамера. (В,С) Схематическая иллюстрация двухступенчатой стратегии хроматографии, показанная для (B) гексамеров септина человека и (C) октамеров. H обозначает его теги, а S — стрептококки II. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Protocol
Representative Results
Discussion
Метод, описанный здесь, обеспечивает надежную очистку и контроль качества предварительно сформированных гетероолигомеров септина. Некоторые из ключевых вопросов, которые необходимо учитывать для правильного применения метода, заключаются в следующем. Во время стадий элюирования в хроматографических разделениях важно использовать рекомендуемую (или более низкую) скорость потока, чтобы свести к минимуму разбавление септиновых комплексов. Кроме того, чтобы максимизировать восстановление белка на конечной стадии концентрации, колонна концентратора ориентирована таким образом, чтобы раствор не прижимался к фильтру (когда с одной стороны есть только фильтр). Если раствор поступает непосредственно в фильтр, белок прилипает к нему гораздо больше, существенно уменьшая конечный выход. Также важно учитывать, что этап концентрации не всегда необходим. Отбор фракций только из узкого диапазона вокруг пика в хроматограмме обычно дает достаточно высокую концентрацию запаса (>3000 нМ) для многих приложений восстановления (которые обычно работают между 10-300 нМ). Наконец, для контроля качества функциональности септиновых комплексов методом флуоресцентной микроскопии важно правильно пассивировать поверхность микроскопических слайдов, так как септиновые комплексы жадно прилипают к стеклу. Пассивация стеклянных слайдов может осуществляться либо путем функционализации ФАПЧ-ПЭГ, либо путем образования нейтральных (100% DOPC) поддерживаемых липидных бислоев 11,32.
По сравнению с исходным протоколом очистки, впервые описанным в Iv et al.10, наблюдается изменение буферных композиций (таблица 2). КонцентрацияMgCl2 снижена с 5 мМ до 2 мМ, а концентрация и рН Tris-HCl снижены с 50 мМ до 20 мМ и с 8,0 до 7,4 соответственно. Эти изменения были сделаны, чтобы сделать буферные условия совместимыми с исследованиями взаимодействия человеческих септинов с липидными бислоями, актиновыми нитями и микротрубочками 10,11,32. Это связано с тем, что авторы сформировали поддерживаемые липидные бислои и полимеризованный актин в F-буфере, состав которого идентичен составу темного SPB, за исключением присутствия АТФ в F-буфере. Изменение буфера не привело к каким-либо изменениям в качестве или сроке службы очищенных септинов по сравнению с исходными буферами.
Этот способ очищения до сих пор имеет ряд ограничений. Во-первых, различные попытки очистки могут различаться по выходу (0,5-1 мл комплексов септина 2-5 мкМ) и функциональному качеству, что подтверждается способностью образования пучка очищенных комплексов септина. Вот почему очень важно последовательно выполнять проверки качества, описанные в этой статье. Очень хороший контроль времени экспрессии и оптической плотности бактериальной культуры может помочь смягчить разницу в урожайности. Во-вторых, этот очистительный трубопровод не может различать тримеры и гексамеры или тетрамеры и октамеры (рисунок 1B). Тем не менее, эксперименты по контролю качества могут быть использованы для доказательства того, что большинство комплексов септина находятся в своей длинной олигомерной форме. В случае, если требуется еще более узкое распределение размеров олигомеров, хроматография исключения размера может быть вставлена между этапом 1.6. и шаг 1.7. протокола очистки. Этот необязательный шаг, однако, резко снижает урожайность, и он не рекомендуется, если это строго не необходимо. Последнее, более фундаментальное, ограничение связано с использованием E. coli в качестве системы экспрессии для рекомбинантных комплексов септина. Естественно, эта система не допускает посттрансляционных модификаций (ПТМ), о которых сообщалось в клетках животных, таких как фосфорилирование, ацетилирование и сумоилирование 6,51,52,53. Эти посттрансляционные модификации могут быть добавлены путем реализации аналогичной стратегии очистки в клетках насекомых или человека. Кроме того, в этой статье обсуждается только восстановление септинов само по себе, но исследования в клетках показывают, что регуляторные белки, такие как белки из семейства Борга54,55 и анилин 24,25,56, могут оказывать существенное, но плохо изученное влияние на сборку и функции септинов и, следовательно, важны для последующего включения в in vitro. учёба. Протоколов для очистки белков Борга и аниллина было зарегистрировано54,57.
Протокол очистки септина, о котором сообщается здесь, предлагает стандартизированный способ очистки септинов в их олигомерной форме с правильной субъединицей стехиометрии, предлагая важный прогресс по сравнению со многими более ранними исследованиями in vitro, основанными на одиночных субъединицах септина. Несмотря на то, что некоторые септины в определенных контекстах могут действовать как одна субъединица2, современная литература убедительно свидетельствует о том, что в клетках животных септины в основном функционируют в комплексах 9,58. Поэтому использование предварительно сформированных гетероолигомеров, таких как описанные в этой статье и других 10,11,18,32,35,36,37, имеет большое значение для изучения структурных и биофизических свойств септинов in vitro. восстановление для рассечения их функций в клетке. Кроме того, септины являются самособирающимися белками со многими партнерами по взаимодействию, включая мембрану и цитоскелет, что делает их очень интересными для синтетической биологии снизу вверх 59,60,61 и исследований белковых изменений биофизических свойств мембраны, таких как кривизна 42,62,63.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
Мы благодарим Сесилию де Агрела Пинто, Томаса де Гарай и Катарину Хойсерманн за их помощь в экспериментах по массовой фотометрии (iSCAT); Арьен Якоби и Вил Эверс за помощь в работе с ТЕА; Лючию Бальдауф за помощь в работе МДП; Паскаль Вердье-Пинар за советы относительно нативного электрофореза; Агата Шуба и Марджолейн Винкенуг за помощь в организации усилий по очистке септина дрозофилы и визуализации клеток и тканей (PICT-IBiSA), Институт Кюри, член Французской национальной исследовательской инфраструктуры France-BioImaging (ANR10-INBS-04). Это исследование получило финансирование от Нидерландской организации научных исследований (NWO/OCW) через грант «BaSyC-Building a Synthetic Cell» Gravitation (024.003.019) и от Национального агентства по исследованиям (гранты ANR ANR-17-CE13-0014: «SEPTIMORF»; ANR-13-JSV8-0002-01: “SEPTIME”; и ANR-20-CE11-0014-01: “SEPTSCORT”).
Materials
| 488nm laser combiner iLAS2 | Gataca | TIRF microscope | |
| 488nm Sapphire laser lines | Coherent | Confocal microscope | |
| 4k X 4k F416 CMOS camera | TVIPS | For JEM-1400plus | |
| 4x sample buffer nativePAGE | Thermo Fisher scientific | BN2003 | |
| 6-Hydroxy-2,5,7,8-tetramethylchromane-2-carboxylic acid (TROLOX) | Sigma-Aldrich | 238813 | To prevent blinking |
| AKTA pure 25 M1 | GE healthcare | 1680311 | |
| Ampicillin | Sigma-Aldrich | A9518-25G | |
| Carbon Type-B, 300 mesh EM grid | Ted pella | 01813-F | |
| Carbon Type-B, 300 mesh EM grid | Electron micoscopy sciences | CF300-Cu | |
| Cover glass #1.5H | Thorslabs | CG15KH | |
| CSU-X1-M1 confocal unit | Yokogawa | Confocal microscope | |
| Desthiobiotin | Sigma-Aldrich | D1411-1G | |
| Dithiothreitol (DTT) | Sigma-Aldrich | D9779 | |
| DNAse | Sigma-Aldrich | 10104159001 | |
| DOPC | Avanti Polar Lipids | 850375C | |
| Eclipse Ti2-E | Nikon instruments | Confocal microscope | |
| EDTA-free protease inhibtor cocktail | Roche | 481761 | |
| HisTrap HP, 5 mL | GE healthcare | 29-0588-3 | |
| iLAS2 azimuthal TIRF illumination system | Gataca | TIRF microscope | |
| Imidazole | Sigma-Aldrich | 1202-1KG | |
| InstantBlue Protein Gel Stain | Westburg Life Sciences | EP ab119211 | |
| Isopropyl β-D-1-thiogalactopyranoside (IPTG) | Thermo Fisher scientific | 10849040 | |
| iXon Ultra 888 EMCCD camera | Andor | Confocal microscope | |
| iXon Ultra 897 EM-CCD | Andor | TIRF microscope | |
| JEM-1400plus | JOEL | TEM microscope TUDelft | |
| kappa-cassein | Sigma-Aldrich | C0406 | |
| LB broth | Sigma-Aldrich | L3022-6X1KG | |
| Lyzozyme | Sigma-Aldrich | 62971-10G-F | |
| Magnesium Chloride | Sigma-Aldrich | M8266-100G | |
| Magnesium sulfate | Sigma-Aldrich | 746452-1KG | |
| Methylecllulose | Sigma-Aldrich | 8074844 | |
| MilliQ system (Integral 10) | Merck-Millipore | I-water dispenser | |
| Mini protean TGX gels | BIORAD | 4561086 | |
| NativeMark unstained protein standard | Invitrogen | LC0725 | For iSCAT and Native gels |
| NativePAGE 4-16% GELS | Thermo Fisher scientific | BN1002BOX | |
| NativePAGE Running Buffer kit | Thermo Fisher scientific | BN2007 | |
| Nikon Ti2-E | Nikon instruments | TIRF microscope | |
| Nr. 1 Menzel coverslips | Thermo Fisher scientific | 11961988 | |
| parafilm | Sigma-Aldrich | P7668 | |
| Plan Apo ×100/1.45 NA oil immersion objective | Nikon instruments | Confocal microscope | |
| PMSF | Sigma-Aldrich | 10837091001 | |
| Poly(L-lysine)-graft-biotinylated PEG (PLL-PEG) | SuSoS | CHF560.00 | |
| Poly-L-lysine solution 0.01% | Sigma-Aldrich | P4832 | For iSCAT glass slides |
| Pottassium Chloride | Sigma-Aldrich | P9541-1KG | |
| Power supply for native gels | CONSORT | S/N 71638 | |
| POWERPAC UNIVERSAL | BIORAD | 042BR31206 | |
| Protocatechuate 3,4-Dioxygenase (PCD) | Sigma-Aldrich | P8279-25UN | oxygen scavenger – enzyme |
| Protocatechuic acid (PCA) | Sigma-Aldrich | 03930590-50MG | oxygen scavenger – reagent |
| Q500 Sonicator | Qsonica | Q500-110 | |
| Quemesa camera | Olympus | For Tecnai Spirit | |
| Refeyn OneMP | Refeyn | ||
| Sample buffer, laemmli 2x concentrate | Sigma-Aldrich | S3401-10vl | |
| Silicon gaskets | Sigma-Aldrich | GBL103250-10EA | |
| Slide-A-Lyzer Dialysis cassettes 30k MWCO 3mL | Thermo Fisher scientific | 66381 | |
| Spectinomycin | Sigma-Aldrich | PHR1441-1G | |
| StrepTrap HP, 1 mL | GE healthcare | 28-9075-46 | |
| Tecnai Spirit microscope | Thermo Scientific, FEI | TEM microscope Institute Curie | |
| Terrific broth | Sigma-Aldrich | T0918-1KG | |
| Tris/Glyine/SDS buffer | BIORAD | 1610772 | |
| Tris-HCl | Sigma-Aldrich | T5941-1KG | |
| Ultrasonic cleaner | Branson | CPX2800H-E | |
| Vivaspin 6, 30,000 MWCO PES | Sartorius | VS0622 |
Riferimenti
- Mostowy, S., Cossart, P. Septins: The fourth component of the cytoskeleton. Nature Reviews Molecular Cell Biology. 13 (3), 183-194 (2012).
- Shuman, B., Momany, M. Septins from protists to people. Frontiers in Cell and Developmental Biology. 9, 3802 (2022).
- Bridges, A. A., Gladfelter, A. S. Septin form and function at the cell cortex. Journal of Biological Chemistry. 290 (28), 17173-17180 (2015).
- Smith, C., et al. Septin 9 exhibits polymorphic binding to F-actin and inhibits myosin and cofilin activity. Journal of Molecular Biology. 427 (20), 3273-3284 (2015).
- Gilden, J. K., Peck, S., Chen, Y. C. M., Krummel, M. F. The septin cytoskeleton facilitates membrane retraction during motility and blebbing. Journal of Cell Biology. 196 (1), 103-114 (2012).
- Marquardt, J., Chen, X., Bi, E. Architecture, remodeling, and functions of the septin cytoskeleton. Cytoskeleton. 76 (1), 7-14 (2018).
- Van Ngo, H., Mostowy, S. Role of septins in microbial infection. Journal of Cell Science. 132 (9), (2019).
- Fung, K. Y. Y., Dai, L., Trimble, W. S. Cell and molecular biology of septins. International Review of Cell and Molecular Biology. 310, 289-339 (2014).
- Kinoshita, M., Field, C. M., Coughlin, M. L., Straight, A. F., Mitchison, T. J. Self- and actin-templated assembly of mammalian septins. Developmental Cell. 3 (6), 791-802 (2002).
- Iv, F., et al. Insights into animal septins using recombinant human septin octamers 2 with distinct SEPT9 isoforms. Journal of Cell Science. 134 (15), (2021).
- Szuba, A., et al. Membrane binding controls ordered self-assembly of animal septins. eLife. 10, 63349 (2021).
- Kinoshita, M. Assembly of mammalian septins. Journal of Biochemistry. 134 (4), 491-496 (2003).
- Connolly, D., et al. Septin 9 isoform expression, localization and epigenetic changes during human and mouse breast cancer progression. Breast Cancer Research. 13 (4), 76 (2011).
- Connolly, D., et al. Septin 9 amplification and isoform-specific expression in peritumoral and tumor breast tissue. Biological Chemistry. 395 (2), 157-167 (2014).
- Estey, M. P., Di Ciano-Oliveira, C., Froese, C. D., Bejide, M. T., Trimble, W. S. Distinct roles of septins in cytokinesis: SEPT9 mediates midbody abscission. Journal of Cell Biology. 191 (4), 741-749 (2010).
- John, C. M., et al. The Caenorhabditis elegans septin complex is nonpolar. EMBO Journal. 26 (14), 3296-3307 (2007).
- Field, C. M., et al. A purified Drosophila septin complex forms filaments and exhibits GTPase activity. Journal of Cell Biology. 133 (3), 605-616 (1996).
- Bertin, A., et al. Saccharomyces cerevisiae septins: Supramolecular organization of heterooligomers and the mechanism of filament assembly. Proceedings of the National Academy of Sciences of the United States of America. 105 (24), 8274-8279 (2008).
- Sellin, M. E., Sandblad, L., Stenmark, S., Gullberg, M. Deciphering the rules governing assembly order of mammalian septin complexes. Molecular Biology of the Cell. 22 (17), 3152-3164 (2011).
- Akil, A., et al. Septin 9 induces lipid droplets growth by a phosphatidylinositol-5-phosphate and microtubule-dependent mechanism hijacked by HCV. Nature Communications. 7, 12203 (2016).
- Tanaka-Takiguchi, Y., Kinoshita, M., Takiguchi, K. Septin-mediated uniform bracing of phospholipid membranes. Current Biology. 19 (2), 140-145 (2009).
- Omrane, M., et al. Septin 9 has two polybasic domains critical to septin filament assembly and Golgi integrity. iScience. 13, 138-153 (2019).
- Carim, S. C., Kechad, A., Hickson, G. R. X. Animal cell cytokinesis: The rho-dependent actomyosin-anilloseptin contractile ring as a membrane microdomain gathering, compressing, and sorting machine. Frontiers in Cell and Developmental Biology. 8, 575226 (2020).
- El Amine, N., Kechad, A., Jananji, S., Hickson, G. R. X. Opposing actions of septins and Sticky on Anillin promote the transition from contractile to midbody ring. Journal of Cell Biology. 203 (3), 487-504 (2013).
- Renshaw, M. J., Liu, J., Lavoie, B. D., Wilde, A. Anillin-dependent organization of septin filaments promotes intercellular bridge elongation and Chmp4B targeting to the abscission site. Open Biology. 4 (1), 130190 (2014).
- Vogt, E. T., et al. The ultrastructural organization of actin and myosin II filaments in the contractile ring: new support for an old model of cytokinesis. Molecular Biology of the Cell. 28 (5), 613-623 (2017).
- Mavrakis, M., et al. Septins promote F-actin ring formation by crosslinking actin filaments into curved bundles. Nature Cell Biology. 16 (4), 322-334 (2014).
- Karasmanis, E. P., et al. A septin double ring controls the spatiotemporal organization of the ESCRT machinery in cytokinetic abscission. Current Biology. 29 (13), 2174-2182 (2019).
- Hagiwara, A., et al. Submembranous septins as relatively stable components of actin-based membrane skeleton. Cytoskeleton. 68 (9), 512-525 (2011).
- Calvo, F., et al. Cdc42EP3/BORG2 and septin network enables mechano-transduction and the emergence of cancer-associated fibroblasts. Cell Reports. 13 (12), 2699-2714 (2015).
- Salameh, J., Cantaloube, I., Benoit, B., Poüs, C., Baillet, A. Cdc42 and its BORG2 and BORG3 effectors control the subcellular localization of septins between actin stress fibers and microtubules. Current Biology. 31 (18), 4088-4103 (2021).
- Kuzmić, M., et al. Septin-microtubule association via a motif unique to isoform 1 of septin 9 tunes stress fibers. Journal of Cell Science. 135 (1), (2022).
- Shindo, A., et al. Septin-dependent remodeling of cortical microtubule drives cell reshaping during epithelial wound healing. Journal of Cell Science. 131 (12), (2018).
- Hu, Q., Nelson, W. J., Spiliotis, E. T. Forchlorfenuron alters mammalian septin assembly, organization, and dynamics. Journal of Biological Chemistry. 283 (43), 29563-29571 (2008).
- Mavrakis, M., Tsai, F. C., Koenderink, G. H. Purification of recombinant human and Drosophila septin hexamers for TIRF assays of actin-septin filament assembly. Methods in Cell Biology. 136, 199-220 (2016).
- Nakos, K., Radler, M. R., Spiliotis, E. T. Septin 2/6/7 complexes tune microtubule plus-end growth and EB1 binding in a concentration- and filament-dependent manner. Molecular Biology of the Cell. 30 (23), 2913-2928 (2019).
- Kaplan, C., et al. Absolute arrangement of subunits in cytoskeletal septin filaments in cells measured by fluorescence microscopy. Nano Letters. 15 (6), 3859-3864 (2015).
- Castro, D. K. S. V., et al. A complete compendium of crystal structures for the human SEPT3 subgroup reveals functional plasticity at a specific septin interface. IUCrJ. 7, 462-479 (2020).
- Jiao, F., Cannon, K. S., Lin, Y. -. C., Gladfelter, A. S., Scheuring, S. The hierarchical assembly of septins revealed by high-speed AFM. Nature Communications. 11 (1), 1-13 (2020).
- Bertin, A., et al. Phosphatidylinositol-4,5-bisphosphate promotes budding yeast septin filament assembly and organization. Journal of Molecular Biology. 404 (4), 711-731 (2010).
- Bridges, A. A., Jentzsch, M. S., Oakes, P. W., Occhipinti, P., Gladfelter, A. S. Micron-scale plasma membrane curvature is recognized by the septin cytoskeleton. Journal of Cell Biology. 213 (1), 23-32 (2016).
- Beber, A., et al. Membrane reshaping by micrometric curvature sensitive septin filaments. Nature Communications. 10, 420 (2019).
- Zhou, R., Shi, Y., Yang, G. Expression, purification, and enzymatic characterization of intramembrane proteases. Methods in Enzymology. 584, 127-155 (2017).
- Diebold, M. L., Fribourg, S., Koch, M., Metzger, T., Romier, C. Deciphering correct strategies for multiprotein complex assembly by co-expression: Application to complexes as large as the histone octamer. Journal of Structural Biology. 175 (2), 178-188 (2011).
- Lebedeva, M. A., Palmieri, E., Kukura, P., Fletcher, S. P. Emergence and rearrangement of dynamic supramolecular aggregates visualized by interferometric scattering microscopy. ACS Nano. 14 (9), 11160-11168 (2020).
- Ludtke, S. J., Baldwin, P. R., Chiu, W. EMAN: Semiautomated software for high-resolution single-particle reconstructions. Journal of Structural Biology. 128 (1), 82-97 (1999).
- Zivanov, J., et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife. 7, 42166 (2018).
- Frank, J., et al. SPIDER and WEB: Processing and visualization of images in 3D electron microscopy and related fields. Journal of Structural Biology. 116 (1), 190-199 (1996).
- Young, G., Kukura, P. Interferometric scattering microscopy. Annual Review of Physical Chemistry. 70, 301-322 (2019).
- Young, G., et al. Quantitative mass imaging of single biological macromolecules. Science. 360 (6387), 423-427 (2018).
- Hernández-Rodríguez, Y., Momany, M. Posttranslational modifications and assembly of septin heteropolymers and higher-order structures. Current Opinion in Microbiology. 15 (6), 660-668 (2012).
- Ribet, D., et al. SUMOylation of human septins is critical for septin filament bundling and cytokinesis. Journal of Cell Biology. 216 (12), 4041-4052 (2017).
- Sinha, I., et al. Cyclin-dependent kinases control septin phosphorylation in Candida albicans hyphal development. Developmental Cell. 13 (3), 421-432 (2007).
- Sheffield, P. J., et al. Borg/Septin interactions and the assembly of mammalian septin heterodimers, trimers, and filaments. Journal of Biological Chemistry. 278 (5), 3483-3488 (2003).
- Joberty, G., et al. Borg proteins control septin organization and are negatively regulated by Cdc42. Nature Cell Biology. 3 (10), 861-866 (2001).
- Chen, X., Wang, K., Svitkina, T., Bi, E. Critical roles of a RhoGEF-anillin module in septin architectural remodeling during cytokinesis. Current Biology. 30 (8), 1477-1490 (2020).
- Kučera, O., et al. Anillin propels myosin-independent constriction of actin rings. Nature Communications. 12 (1), 1-12 (2021).
- Hsu, S. C., et al. Subunit composition, protein interactions, and structures of the mammalian brain sec6/8 complex and septin filaments. Neuron. 20 (6), 1111-1122 (1998).
- Olivi, L., et al. Towards a synthetic cell cycle. Nature Communications. 12 (1), 1-11 (2021).
- Hürtgen, D., Härtel, T., Murray, S. M., Sourjik, V., Schwille, P. Functional modules of minimal cell division for synthetic biology. Advanced Biosystems. 3 (6), 1800315 (2019).
- Jia, H., Schwille, P. Bottom-up synthetic biology: Reconstitution in space and time. Current Opinion in Biotechnology. 60, 179-187 (2019).
- Cannon, K. S., Woods, B. L., Crutchley, J. M., Gladfelter, A. S. An amphipathic helix enables septins to sense micrometer-scale membrane curvature. The Journal of Cell Biology. 218 (4), 1128-1137 (2019).
- Lobato-Márquez, D., Mostowy, S. Septins recognize micron-scale membrane curvature. Journal of Cell Biology. 213 (1), 5-6 (2016).
