Le cellule staminali adulte (SCS) sono essenziali per mantenere l'omeostasi tissutale, sostituendo le cellule che muoiono e riparare tessuti danneggiati su lesioni. Questi SC sono definite dalla loro capacità di subire un continuo auto-rinnovamento e di differenziarsi in varie linee cellulari 1-3. I sistemi più studiati, che dipendono SCs adulti per il rifornimento, includono il sistema ematopoietico, l'intestino ed il 1,2,4 pelle.
Durante l'embriogenesi, la pelle inizia come un singolo strato di cellule epidermiche. Morfogenesi del follicolo pilifero (HF) inizia quando le cellule mesenchimali popolano la pelle e formano un sottostante derma collagene 5. Specializzato cellule mesenchimali, che poi costituiscono la papilla dermica (DP), organizzano direttamente sotto lo strato epidermico e stimolano l'epitelio di formare placodi capelli che iniziano a crescere verso il basso 6. Altamente cellule proliferanti della matrice, situato nella parte inferiore del HF,busta queste cellule mesenchimali e formare il bulbo pilifero, mentre lo strato interno comincia a differenziarsi in cilindri concentrici per formare il fusto del capello (HS) e nelle vicinanze della guaina interna della radice (IRS) 2,3.
Nella vita postnatale l'epidermide della pelle è composta da tre vani: l'epidermide interfollicolare (IFE), la ghiandola sebacea (SG) e l'HF. In contrasto con la IFE e SG che sono in un costante stato di omeostasi, l'HF è un mini-organo dinamico che subisce continui cicli di crescita (anagen), la distruzione (catagen) e di riposo (telogen) 4,7. Le cellule staminali del follicolo pilifero (HFSCs) che il carburante questo ciclo perpetuo, risiedono in una nicchia specializzata all'interno della HF conosciuto come il rigonfiamento 4. Durante anagen i HFSCs uscire dal rigonfiamento, a seguito di segnali di attivazione dal DP, iniziare proliferare e scendere verso il basso creando così una lunga scia lineare di cellule conosciute come la guaina esterna radice (ORS) 8-10. Le celle di matrice, checircondare il DP alla base della HF, rapidamente ciclo e migrare verso l'alto subire differenziazione terminale generando l'HS e l'IRS 10 (Figura 1). La durata di anagen determina la lunghezza dei capelli e dipende dalla capacità proliferativa e la differenziazione delle cellule della matrice 6. Quando l'HF entra catagen, le celle di matrice transito di amplificazione nel bulbo cessano di proliferare, apoptosi e regrediscono completamente tirando la DP verso l'alto fino a raggiungere la parte non bicicletta della HF 8,11. Durante questa retrazione HF forma una struttura temporanea noto come il filamento epiteliale, che è caratteristica di catagen, e contiene molte cellule apoptotiche. Nei topi, catagen dura tra 3-4 giorni ed è altamente sincronizzato nel primo ciclo del capello. Quando la HF raggiunge telogen tutti HFSCs diventano quiescenti. Le fasi distinte del ciclo HF sono inoltre caratterizzati da cambiamenti nel colore della pelle del topo causa mla produzione elanin. I cambiamenti della pelle dal nero durante anagen al grigio scuro durante catagen al rosa durante telogen 6,7,12,13.
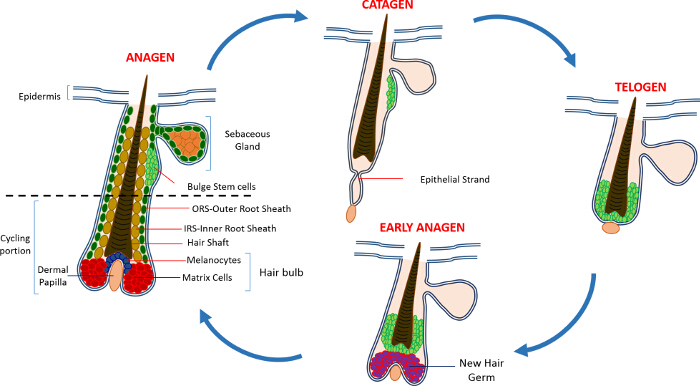
Figura 1: Il ciclo dei capelli follicolo. Il HF è composto da una parte superiore permanente e una porzione ciclismo inferiore costantemente rimodellamento che subisce continui cicli di rapida crescita (anagen), la distruzione (catagen) e una fase di quiescenza parente o di riposo (telogen). Clicca qui per visualizzare un più grande versione di questa figura.
SCS mantenendo la HF sono stati inizialmente identificati utilizzando esperimenti Chase, con timidina triziata, che ha rivelato una popolazione di cellule di sostegno etichette ciclismo lento (LRC) che risiedevano nella regione permanente del HF appena sotto la SG 14. I progressi nella HFSCCaratterizzazione rivelato un piccolo numero di marcatori che possono essere utilizzati per identificare e isolare SCs specifici dalla nicchia HF 15. Forse il miglior marcatore per l'arricchimento di HFSCs è CD34, un marker superficie cellulare anche identificato come un marcatore SC ematopoietiche negli esseri umani 16. All'interno di questo CD34 + popolazioni due distinte popolazioni sono stati anche isolati sulla base di espressione α6 integrina 2. Un altro indicatore è la cheratina 15 (K15), che è altamente espresso nella regione bulge, co-localizza con espressione CD34 e un promotore K15 viene utilizzato per il targeting e isolare HFSCs in animali transgenici 15,17-19. Negli ultimi dieci anni sono stati segnalati anche diversi altri distinte popolazioni di HFSCs e cellule progenitrici di risiedere all'interno del HF 17,20-27.
Una caratteristica interessante aggiuntiva di HFSCs è il loro contributo alla riparazione della pelle. In condizioni normali HFSCs ricostituire l'HF e non prendono parte a IFE omeostasi. However, in risposta a ferita, queste cellule uscire la loro nicchia SC e di aiuto nel ripopolare l'IFE 9. Abbiamo recentemente dimostrato che i topi cancellati per il display gene Sept4 / ARTS pro-apoptotica un aumento del numero di CD34, K15 e Sox9 + HFSCs, che dimostrano una resistenza all'apoptosi. HFSCs sono stati isolati da Sept4 / ARTS – / – pelli dorsali utilizzando la fluorescenza delle cellule attivate (FACS) e c'era più di due volte maggiore del numero di cellule CD34 + e K15 + HFSCs. Questi Sept4 / arte – / – HFSCs sono state espanse in vitro e non hanno dato luogo a solo più colonie, ma sono stati anche in grado di resistere a condizioni più severe rispetto ai controlli 28.
Come risultato di avere un maggior numero di HFSCs, Sept4 / arte – / – mice guarite significativamente più veloce in risposta alle lesioni asportazione della pelle. Sorprendentemente, Sept4 / arte – / – mice displayeda gran numero di HF rigenerate dal letto della ferita, e cicatrici significativamente più piccoli. Inoltre, i topi cancellati per XIAP (inibitore della X-linked di apoptosi), il target biochimico di ARTI, hanno dimostrato ridotta guarigione 28.
I nostri risultati e il lavoro svolto in altri laboratori hanno dimostrato che HFSCs servire come un modello ideale per lo studio della biologia e della funzione della SCS adulti. Qui, descriviamo la metodologia per l'arricchimento e l'isolamento di HFSCs e cheratinociti epidermici in base alla espressione di quattro marcatori: α6 integrina; β1 integrine; Sca-1 (un marker per cheratinociti epidermici) e CD34. Isolamento analogo di K15 + HFSCs può essere eseguita anche con il giornalista del mouse K15-GFP 19.
