Yetişkin kök hücreleri (SC'ler) ölen hücrelerin yerine doku vücut dengesinin sürdürülmesinde ve yaralanma üzerine hasarlı dokuların onarımı için gereklidir. Bu SC'ler sürekli kendini yenileme geçmesi ve çeşitli hücre soyları 1-3 ayırt etme yetenekleri ile tanımlanmıştır. Kendi ikmal için yetişkin SC'ler bağlıdır iyi çalışılan sistemler, hematopoetik sistem, bağırsak ve cilt 1,2,4 bulunmaktadır.
embriyojenez sırasında cilt epidermal hücrelerin tek bir tabaka olarak başlar. Mezenkimal hücreler cilt doldurmak ve altta yatan kollajen dermis 5 oluşturduklarında saç folikülü (HF) morfonogenezi başlar. Aşağıya doğru 6 büyümeye başlar saç placodes oluşturmak için daha sonra, dermal papilla (DP) oluşturan epidermal tabakasının altında doğrudan organize etmek ve epitel teşvik mezenkimal hücreler, uzman. Son derece HF alt kısmında yer alan matris hücreleri, çoğalan,iç katman saç şaft (HS) ve çevresindeki iç kök kılıfı (IRS) 2,3 oluşturmak için konsantrik silindir içine ayırt etmeye başlar ise, bu mezenkimal hücreler zarf ve saç ampul oluştururlar.
interfolliküler epidermis (IFE), yağ bezi (SG) ve HF: doğum sonrası hayatta deri epidermis üç bölmeden oluşmaktadır. Homeostasis sürekli bir durumdadır IFE ve SG aksine, HF sürekli büyüme döngüleri (anagen), yıkım (katagen) ve geri kalanı (telojen) 4,7 uğrar dinamik bir mini organdır. Kıl folikülü kök hücreleri (HFSCs) yakıt bu daimi döngüsü, şişkinlik 4 olarak bilinen HF içinde özel bir niş bulunur. DP etkinleştirme sinyalleri HFSCs şişkinliği çıkış anajen takip sırasında proliferasyon başlar ve aşağı doğru ve böylece dış kök kılıf olarak bilinen hücrelerin uzun lineer iz oluşturma alçalma (ORS) 8-10. Matris hücreleri, yaniHF, hızla döngüsünün üssünde DP'yi çevreleyen ve böylece HS ve IRS 10 (Şekil 1) üreten terminal farklılaşma geçiren yukarı göç. Anagen süresi saç uzunluğunu belirler ve matriks hücrelerinin 6 çoğalma ve farklılaşma kapasitesine bağlıdır. HF katagen girer zaman, ampul ateşkes transit yalıtım matris hücreler çoğalmaya apoptoz geçmesi ve HF 8,11 olmayan bisiklet parçası ulaşıncaya kadar yukarı DP çekerken tamamen gerileme. Bu geri çekme sırasında HF katagen karakteristik epitel iplikçik olarak bilinen geçici bir yapı oluşturur, ve çok sayıda apoptotik hücre içerir. Farelerde, katagen 3-4 arasındaki gün sürer ve son derece ilk saç döngüsünde eşitlenir. HF telojen ulaştığında tüm HFSCs pasif hale gelir. HF döngüsünün farklı aşamaları da m bağlı fare deri rengine değişiklikler ile karakterize edilirelanin üretimi. Katagen sırasında koyu gri anagen sırasında siyah deri değişiklikleri telojen 6,7,12,13 sırasında pembe.
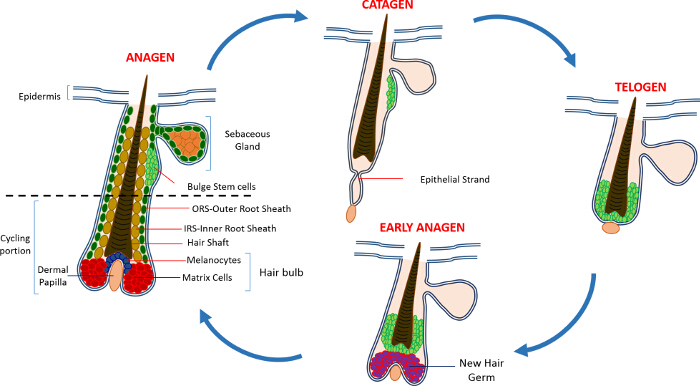
Şekil 1: Saç Folikül Döngüsü. HF kalıcı üst kısmının oluşan ve hızlı büyüme (anagen), yıkım (katagen) ve göreli bir sessizlik fazı veya geri kalanı (telojen) sürekli döngülerine maruz düşük sürekli yeniden yapılanma, bisiklet bölümü. Olan bir büyük görmek için tıklayınız Bu rakamın sürümü.
HF muhafaza SC'ler başlangıçta sadece SG 14 altında HF daimi bölgesinde ikamet yavaş bisiklet etiket istinat hücreler (LRC) nüfusu ortaya öğütülmüş ümidin ile kovalamaca deneyleri, kullanılarak belirlendi. HFSC gelişmelerKarakterizasyon HF niş 15 belirli SC'ler belirlemek ve izole etmek için kullanılabilir belirteçlerin az sayıda tespit edildi. Belki HFSCs zenginleştirilmesi için iyi markör CD34, aynı zamanda, insanlarda 16 bir hematopoietik SC markör olarak tanımlanan bir hücre yüzey markörüdür. Bu CD34 + nüfus içinde iki ayrı nüfus da izole α6 integrin ekspresyonu 2 dayalı olmuştur. Başka bir belirteç CD34 ekspresyonu ile birlikte lokalize ve K-15 promoteri hedefleme ve transgenik hayvanlarda 15,17-19 içinde HFSCs izole edilmesi için kullanılan, keratin yüksek şişiklik bölgesinde ifade edilir 15 (K15) 'dir. Geçtiğimiz on yıl içinde HFSCs ve projenitör hücrelerin birçok farklı popülasyonlar da HF 17,20-27 içinde yer bildirilmiştir.
HFSCs ek heyecan verici özelliği cilt onarım katkıları olduğunu. Normal şartlar altında HFSCs HF doldurmak ve IFE homeostazındaki yer almazlar. However yaralama yanıt olarak, bu hücreler IFE 9 repopulating kendi SC niş ve yardım çıkar. Son zamanlarda fareler pro-apoptotik Sept4 / sanat geni gösterim için apoptoza bir direnç gösterir CD34, 15 K ve SOX9 + HFSCs, artmış sayıda silinmiş göstermiştir. HFSCs Sept4 / sanat izole edilmiştir – / – dorsal derileri Kontrol THP (FACS) kullanılarak CD34 + ve K15 + HFSCs sayısının fazla iki kat daha yüksek olmuştur. Bunlar Sept4 / SANAT – / – HFSCs in vitro genişletilmiş ve değil sadece daha koloniler doğurdu ama kontrollere 28 ile karşılaştırıldığında aynı zamanda sert koşullarına dayanacak şekilde başardık.
HFSCs artan sayısına sahip bir sonucu olarak, Sept4 / sanat – / – farelerinin, deri eksizyon yaralanmaları yanıt olarak önemli ölçüde daha hızlı iyileşti. Çarpıcı, Sept4 / SANAT – / – fareler displayeyara yatağından rejenere HFS ve önemli ölçüde daha küçük izleri da çok sayıda. Ayrıca, XIAP (apoptoz X'e bağlı inhibitörü), SANAT biyokimyasal hedefi için silinen fareler bozulmuş iyileşme 28 gösterdi.
Bizim sonuçları ve diğer laboratuarlarda yapılan çalışma HFSCs yetişkin SC'ler biyolojisi ve fonksiyonu eğitim için ideal bir model olarak hizmet olduğunu göstermiştir. Burada, zenginleştirme ve HFSCs dört markerlerin ekspresyonu göre epidermal keratinositler izolasyonu için bir yöntem açıklanmaktadır: integrin α6; integrin β1; Sca-1 (epidermal keratinositler için bir belirteç) ve CD34. K15 + HFSCs Benzer bir ayrılma K15-GFP raportör fare 19 kullanılarak da gerçekleştirilebilir.
