Les cellules souches adultes (SC) sont essentiels pour maintenir l'homéostasie tissulaire en remplaçant les cellules meurent et la réparation des tissus endommagés lors d'une lésion. Ces SCs sont définis par leur capacité à subir l' auto-renouvellement continu et de se différencier en diverses lignées cellulaires 1-3. Les systèmes les plus étudiés, qui dépendent SCs adultes pour leur reconstitution comprennent le système hématopoïétique, l'intestin et le 1,2,4 cutané.
Au cours de l'embryogenèse, la peau commence par une seule couche de cellules épidermiques. Morphogénèse du follicule pileux (HF) commence lorsque les cellules mésenchymateuses peuplent la peau et forment un derme sous – jacent collagènes 5. Spécialisée des cellules mésenchymateuses, qui constituent la suite de la papille dermique (DP), organisent directement sous la couche de l' épiderme et stimulent l'épithélium pour former placodes cheveux qui commencent à se développer vers le bas 6. Fortement la prolifération des cellules de la matrice, situés à la partie inférieure du HF,envelopper ces cellules mésenchymateuses et forment le bulbe du cheveu, tandis que la couche intérieure commence à se différencier en cylindres concentriques pour former la tige pilaire (HS) et la gaine intérieure entourant la racine (IRS) 2,3.
Dans la vie postnatale l'épiderme de la peau est composé de trois compartiments: l'épiderme interfolliculaires (IFE), la glande sébacée (SG) et le HF. Contrairement à l'IFE et SG qui sont dans un état constant de l' homéostasie, la HF est un mini-organe dynamique qui subit des cycles continus de croissance (anagène de), la destruction (catagène) et de repos (télogène) 4,7. Les cellules souches du follicule pileux (HFSCs) que de carburant ce cycle perpétuel, résident dans un créneau spécialisé dans le HF connu sous le renflement 4. Pendant anagène les HFSCs quitter le renflement, après des signaux d'activation de la DP, commencer à proliférer et descendent vers le bas créant ainsi un long sentier linéaire de cellules connues sous le nom de la gaine externe de la racine (ORS) 8-10. Les cellules de la matrice, quientourer la DP à la base de la HF, le cycle rapidement et migrer vers le haut subissant la différenciation terminale générant ainsi le HS et l'IRS 10 (Figure 1). La durée de la phase anagène détermine la longueur des cheveux et dépend de la capacité de prolifération et la différenciation des cellules de la matrice 6. Lorsque le HF pénètre catagène, les cellules de la matrice de transit-amplification du cessez ampoule de proliférer, subissent l' apoptose et régressent entièrement tout en tirant le DP vers le haut jusqu'à ce qu'il atteigne la partie non-cyclique du HF 8,11. Au cours de cette rétraction HF forme une structure temporaire connu sous le brin de l'épithélium, qui est caractéristique de la phase catagène et contient de nombreuses cellules apoptotiques. Chez la souris, catagène dure entre 3-4 jours et est hautement synchronisé dans le premier cycle pilaire. Lorsque le HF atteint télogène tous HFSCs deviennent repos. Les différentes étapes du cycle HF sont également caractérisés par des changements dans la couleur de la peau de la souris, du fait de mproduction elanin. La peau passe du noir au cours de anagène au gris foncé pendant catagène au rose pendant télogène 6,7,12,13.
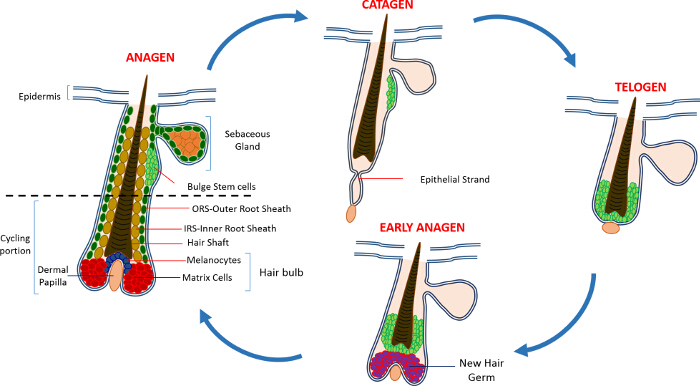
Figure 1: Le cycle follicule pileux. Le HF est composé d'une partie permanente supérieure et, une partie de cyclisme remodelage inférieure constante qui subit des cycles continus de croissance rapide (anagène), la destruction (catagène) et une phase de quiescence relative ou de repos (télogène). S'il vous plaît cliquer ici pour voir une plus grande version de ce chiffre.
Les SCs maintenant la HF ont été initialement identifiés en utilisant des expériences de chasse, avec de la thymidine tritiée, qui a révélé une population de cellules d'étiquettes cycliste lente retenue (LRC) qui résidaient dans la région permanente du HF juste en dessous du SG 14. Advances in HFSCla caractérisation a révélé un petit nombre de marqueurs qui peuvent être utilisés pour identifier et isoler SCs spécifiques de la niche HF 15. Peut-être le meilleur marqueur pour l' enrichissement de HFSCs est CD34, un marqueur de surface cellulaire a également identifié comme un marqueur SC 16 hématopoïétique chez l' homme. Dans ce CD34 + populations deux populations distinctes ont également été isolés sur la base de α6 intégrine expression 2. Un autre marqueur est la kératine 15 (K15) , qui est fortement exprimé dans la région de la bosse, la co-localise avec l' expression du CD34 et un promoteur K15 est utilisé pour le ciblage et l' isolement HFSCs chez des animaux transgéniques 15,17-19. Dans la dernière décennie , plusieurs autres populations distinctes de HFSCs et les cellules progénitrices ont également été signalés à résider dans le HF 17,20-27.
Une caractéristique supplémentaire intéressante de HFSCs est leur contribution à la réparation de la peau. Dans des conditions normales HFSCs reconstituent le HF et ne prennent pas part à IFE homéostasie. However, en réponse à la blessure, ces cellules quitter leur niche de SC et de l' aide à repeupler la IFE 9. Nous avons récemment démontré que les souris supprimés pour l'affichage du gène Sept4 / ARTS pro-apoptotique un nombre accru de CD34, K15 et Sox9 + HFSCs, qui démontrent une résistance à l' apoptose. HFSCs ont été isolés de Sept4 / ARTS – / – peaux dorsales utilisent cellules activées par fluorescence (FACS) et il y avait plus de deux fois plus dans le nombre de CD34 + et K15 + HFSCs. Ces Sept4 / ARTS – / – HFSCs ont été élargis in vitro et non seulement ont donné lieu à plus de colonies , mais sont également capables de résister à des conditions plus sévères par rapport aux témoins 28.
En conséquence d'avoir un nombre accru de HFSCs, Sept4 / ARTS – / – souris guéries nettement plus rapide en réponse à des blessures d'excision de la peau. Il est frappant, Sept4 / ARTS – / – souris displayeda grand nombre de formations sanitaires régénérées du lit de la plaie, et significativement plus petites cicatrices. En outre, les souris supprimées pour XIAP (inhibiteur lié à l'X de l' apoptose), la cible biochimique des ARTS, ont démontré une mauvaise cicatrisation 28.
Nos résultats et les travaux effectués dans d'autres laboratoires ont montré que HFSCs servent comme un modèle idéal pour étudier la biologie et la fonction de SCs adultes. Ici, nous décrivons la méthode utilisée pour l'enrichissement et l'isolement de HFSCs et kératinocytes de l'épiderme sur la base de l'expression de quatre marqueurs: α6 intégrine; intégrine β1; Sca-1 (un marqueur pour les kératinocytes de l'épiderme) et CD34. Isolement similaire de K15 + HFSCs peut également être effectuée en utilisant le K15-rapporteur GFP 19 souris.
