Stem Embedding and Sectioning:
The use of the homemade plastic mold to embed the stems in 7% agarose proved to be fast and easy (Figure 1). The two parts (A and B; Figure 1) of the embedded vial system make it simple to easily release the stem embedded in agarose as the agarose does not stick to the vial parts that are inert, keeping the system clean. The vials can be reused multiple times for years. The convenience of storing the embedded stem also makes the next steps easier. In order to save sometime, similar embedded stems can be grouped (up to 3) on a single sectioning plate for sectioning.
Observation of Aromatics Under UV:
The stem sections were mounted in distilled water on to the glass slides and observed under UV light. The aromatic compounds, including the lignin in the cells, can be visualized by their autofluorescence under UV light. Xylem (Xy) and interfascicular fibers (Fi) showed autofluorescence under UV illumination but not in pith (Pi) and cortex or epidermis (Ep) (Figure 2), because lignin, an aromatic polymer, is deposited during secondary cell-wall biosynthesis. In some instances, when plants accumulate significant amounts of aromatics in their vacuoles (e.g., anthocyanin), fluorescence can be observed in cortical cells (data not shown). Stem cross-sections were analyzed under 5X, 10X, 20X, and 40X magnification (Figure 2 Panels A, B, C, and D–E, respectively) to better visualize cell wall aromatic distribution across the stem.
Observation of Lignin in the Stem Sections Stained with Phloroglucinol and Mäule Stains:
The xylem composed of vessels and fibers and interfascicular fibers were the only elements of the stem that were stained by the lignin stains (phloroglucinol and Mäule) using a wild type stem section sample. Phloroglucinol stain reacts with cinnamaldehyde end-groups of lignin to give a pink or fuchsia color4 (Figure 3). As observed under UV illumination, a fuchsia coloration is observed in xylem and interfascicular fibers but is absent in pith and cortex or epidermis (Figure 3). Stem cross-sections were analyzed under 5X, 10X, 20X, and 40X magnification (Figure 3 Panels A, B, C, and D–E, respectively) to better visualize phloroglucinol stain distribution across the stem and differentiate the major tissues; xylem and interfascicular fibers; pith and cortex; and epidermis. Usually the intensity of the color correlates with the level of lignification in a qualitative manner — except when the analyzed mutant or transgenic is abnormally enriched in cinnamaldehyde-derived units (e.g. cad mutant)14. The Mäule stain is specific in detecting the syringyl lignin units in xylem and interfascicular fibers. Red coloration indicates the presence of syringyl lignin units in the lignin elements (Figure 4)18. However, a brighter red coloration is observed in the fibers when compared to the xylem tissues which suggests that the fibers contain a higher level of S lignin whereas the xylem is more enriched in G units. As observed after phloroglucinol staining, a red coloration is observed in xylem and interfascicular fibers but is absent in pith and cortex or epidermis (Figure 4). It also correlates with the lignin distribution observed under UV (Figure 2) and shows that syringyl units are present in every lignified tissue of this stem sample. Stem cross-sections were analyzed under 5X, 10X, 20X, and 40X magnification (Figure 2 Panels A, B, C, and D–E, respectively) to better visualize Mäule stain distribution across the stem and differentiate the major tissues: xylem and interfascicular fibers, pith and cortex, and epidermis. It is important to note that the amount and distribution of syringyl units between lignified tissues vary with the age of the stem and can be absent in a young stem (data not shown).
Observation of Stem Sections Stained with Calcofluor White and Congo Red Stains:
Calcofluor white stains cellulose, callose, and other nonsubstituted or weakly substituted β-glucans; whereas congo red stains directly to β-(1→4)-glucans and particularly to cellulose. Stem sections stained with calcofluor white were observed under UV light6-9. Congo red-stained sections were observed under blue-light excitation using a filter with a bandpass of 560/40. Both calcofluor white and congo red stained the epidermis, cortex, and pith; this is because all of these tissues contain cellulose as major polysaccharide polymer in their cell walls (Figure 5 and Figure 6, respectively). In contrast to calcofluor white, congo red stained polysaccharides better in the xylem and interfascicular fibers and seems less affected by the presence of lignin (Figure 5 and Figure 6)10,11. Stem cross-sections were analyzed under 5X, 10X, 20X, and 40X magnification (Panels A, B, C, and D-E, respectively of Figures 5 and 6) to better visualize calcofluor white and congo red stain distributions across the stem and the major tissues (xylem and interfascicular fibers, pith and cortex, and epidermis).
Observation of Stem Sections Stained with Toluidine Blue O:
Toluidine blue O is classified as a polychromatic dye because it reacts with different chemical components of cells differently and results in a multi-colored specimen (Figure 7). The colors generated can provide information on the nature of the cell and its walls. Toluidine blue O is a cationic dye that binds to negatively charged groups5. An aqueous solution of this dye is blue, but different colors are generated when the dye binds with different anionic groups in the cell6,9. For example, a pinkish purple color will appear when the dye reacts with carboxylated polysaccharides such as pectic acid; green, greenish blue, or bright blue with poly-aromatic substances such as lignin and tannins; and purplish or greenish blue with nucleic acids. Toluidine blue O of stem cross-sections revealed that xylem and interfascicular fibers are lignified since they show a greenish blue or blue coloration (Figure 7, Panel C), which is in agreement with the UV and Ph-HCl staining observations (Figures 2 and 3). In contrast, the pith and cortex and epidermis tissues show greenish blue or blue coloration because, although they are not lignified, they contain some pectin polymers in their cell walls. Stem cross-sections were analyzed under 5X, 10X, 20X, and 40X magnification (Figure 7 Panels A, B, C, and D–E, respectively) to better visualize Toluidine blue O stain distribution across the stem and to differentiate the major tissues.

Figure 1. Homemade mold for embedding the stems. On the left side; top of the 2 ml screw-cap microcentrifuge tube (Part A) and the bottom from the 0.6 ml microcentrifuge tube (Part B). On the right side; the Parafilm holding the two parts; A and B to form the final mold.
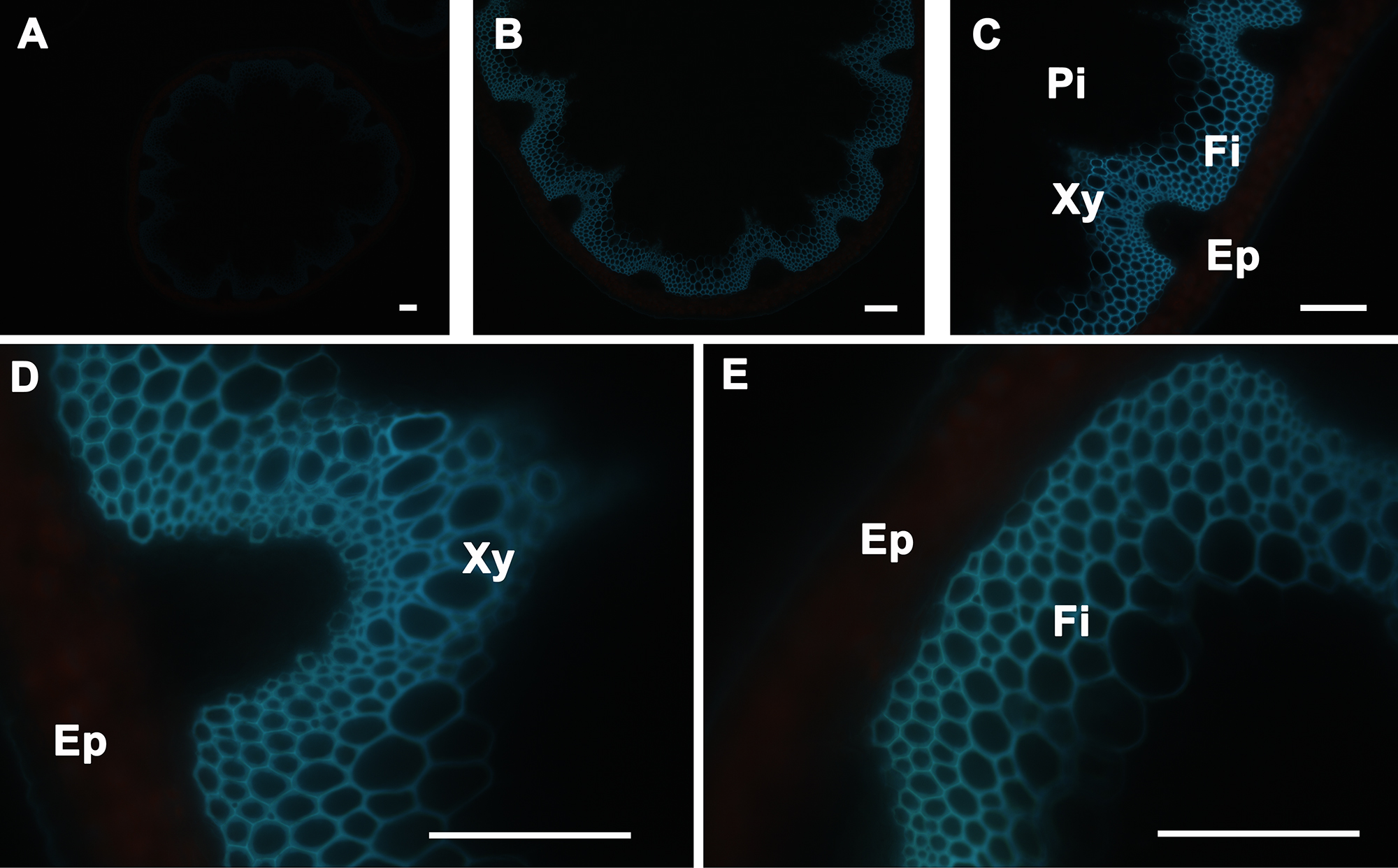
Figure 2. UV Fluorescence of A. thaliana stem cross-sections. Transverse sections of a wild type A. thaliana stem under UV light fluorescence showing the presence of aromatic compounds, including lignin, only in the walls of interfascicular fibers and xylem cells (A–E). The positions of the epidermis and cortex (Ep), interfascicular fibers (Fi), pith (Pi), and xylem (Xy) are indicated. Magnifications: Panel A: 5X; Panel B: 10X; Panel C: 20X; Panels D and E: 40X. Bar = 100 µm.
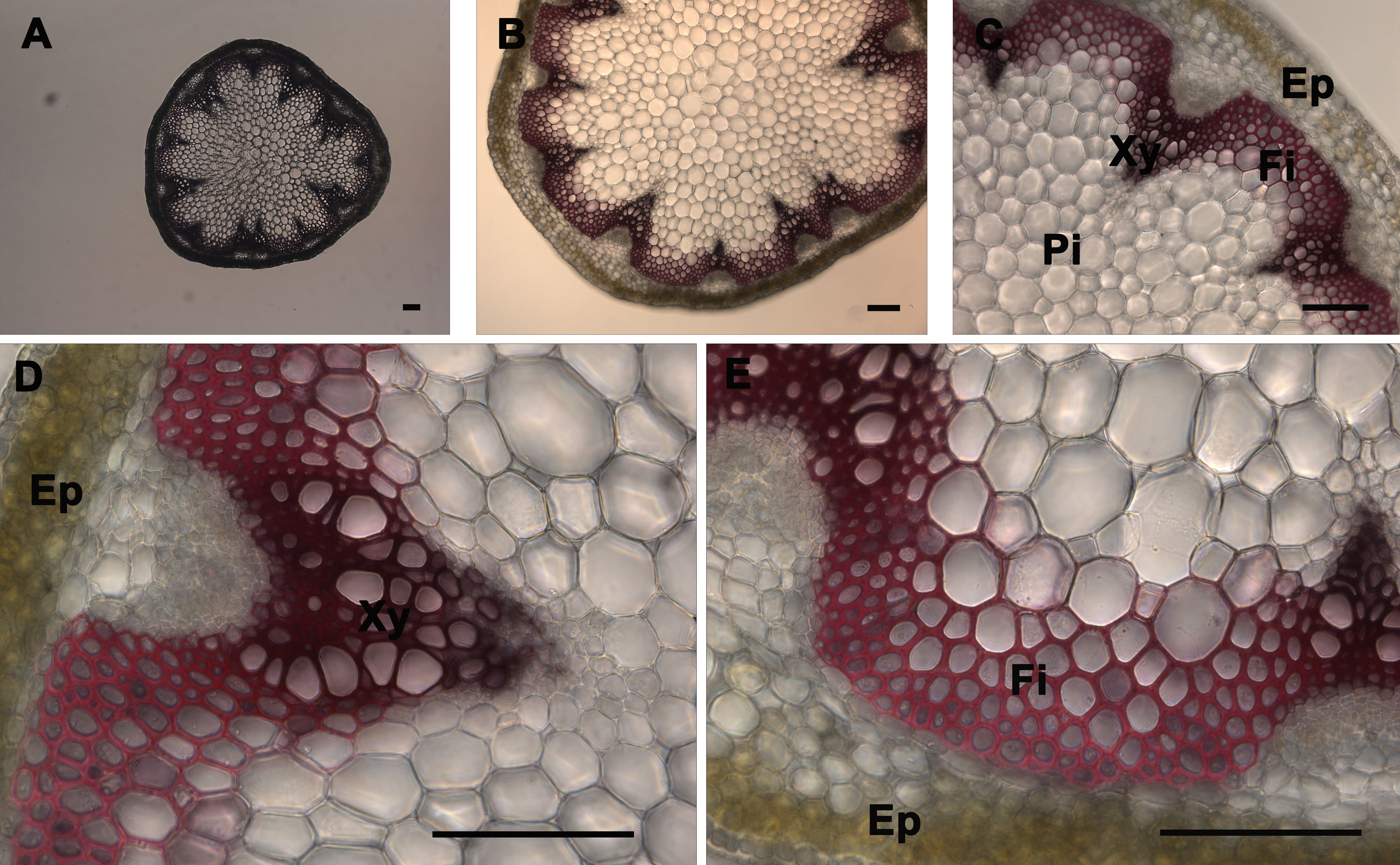
Figure 3. Phloroglucinol-HCl staining of A. thaliana stem cross-sections. Phloroglucinol-HCl staining (pink or fuchsia color) of a wild type A. thaliana stem section showing the normal lignin deposition in the walls of interfascicular fibers and xylem cells (A–E). The positions of the epidermis and cortex (Ep), interfascicular fibers (Fi), pith (Pi), and xylem (Xy) are indicated. Magnifications: Panel A: 5X; Panel B: 10X; Panel C: 20X; Panels D and E: 40X. Bar = 100 µm.
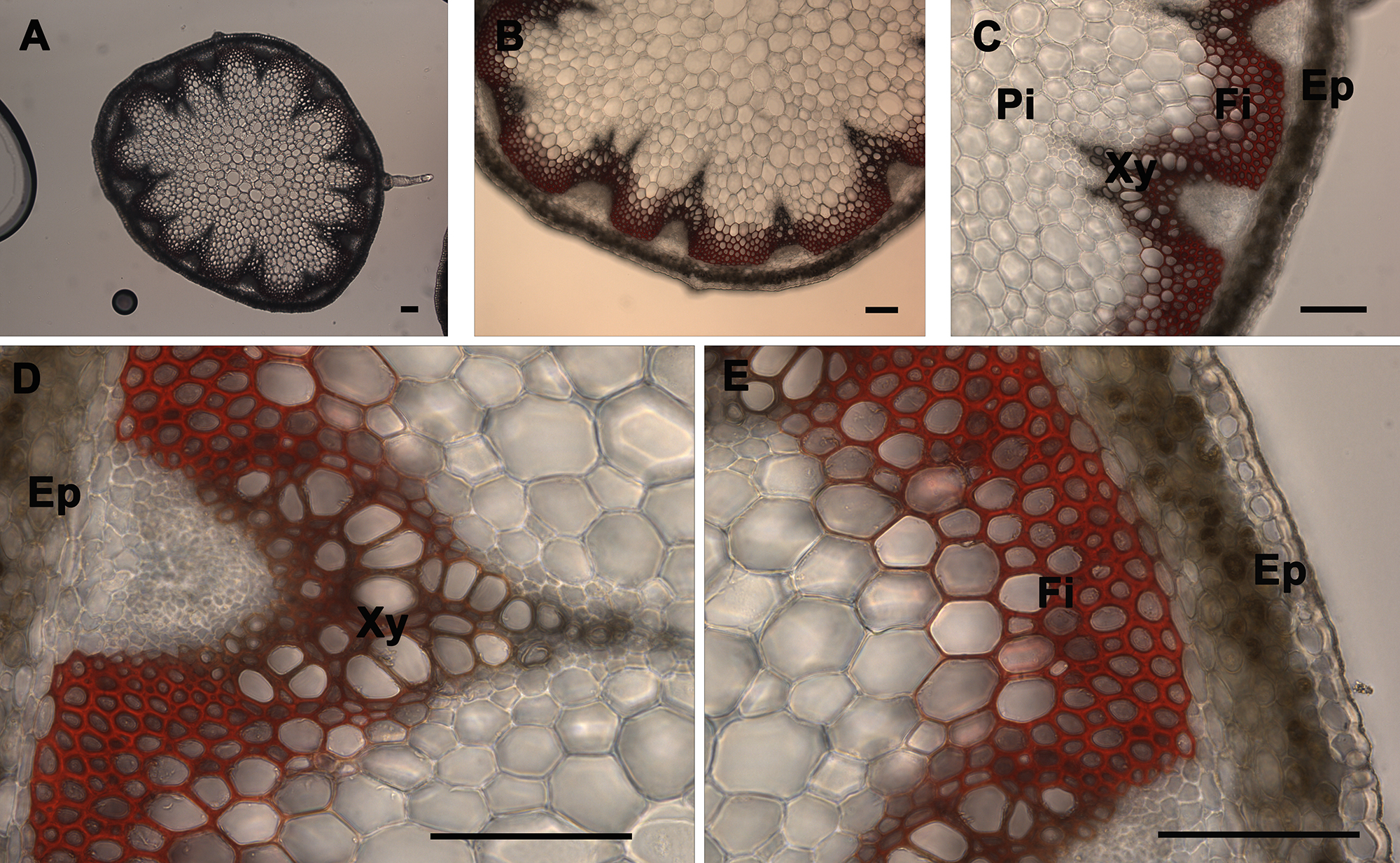
Figure 4. Mäule staining of A. thaliana stem cross-sections. Mäule staining (red color) of a wild type A. thaliana stem section showing the normal lignin deposition in the walls of interfascicular fibers and xylem cells (A–E). The positions of the epidermis and cortex (Ep), interfascicular fibers (Fi), pith (Pi), and xylem (Xy) are indicated. Magnifications: Panel A: 5X; Panel B: 10X; Panel C: 20X; Panels D and E: 40X. Bar = 100 µm.
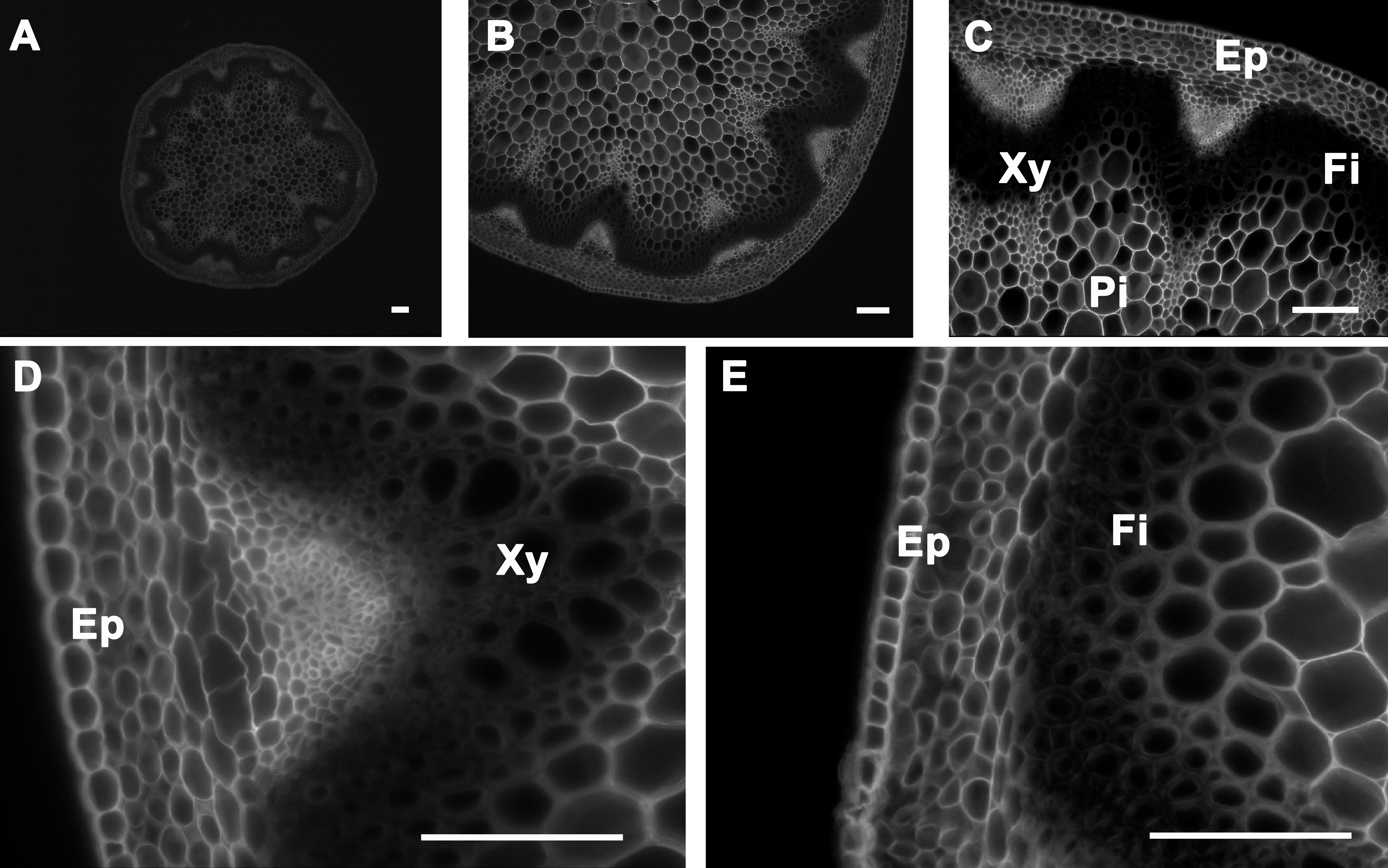
Figure 5. Calcofluor white staining of A. thaliana stem cross-sections. Calcofluor white staining of a wild type A. thaliana stem section showing cellulose deposition in the cell walls of cells of the epidermis, cortex, pith, interfascicular fibers, and xylem (A–E). The positions of the epidermis and cortex (Ep), interfascicular fibers (Fi), pith (Pi), and xylem (Xy) are indicated. Magnifications: Panel A: 5X; Panel B: 10X; Panel C: 20X; Panels D and E: 40X. Bar = 100 µm.
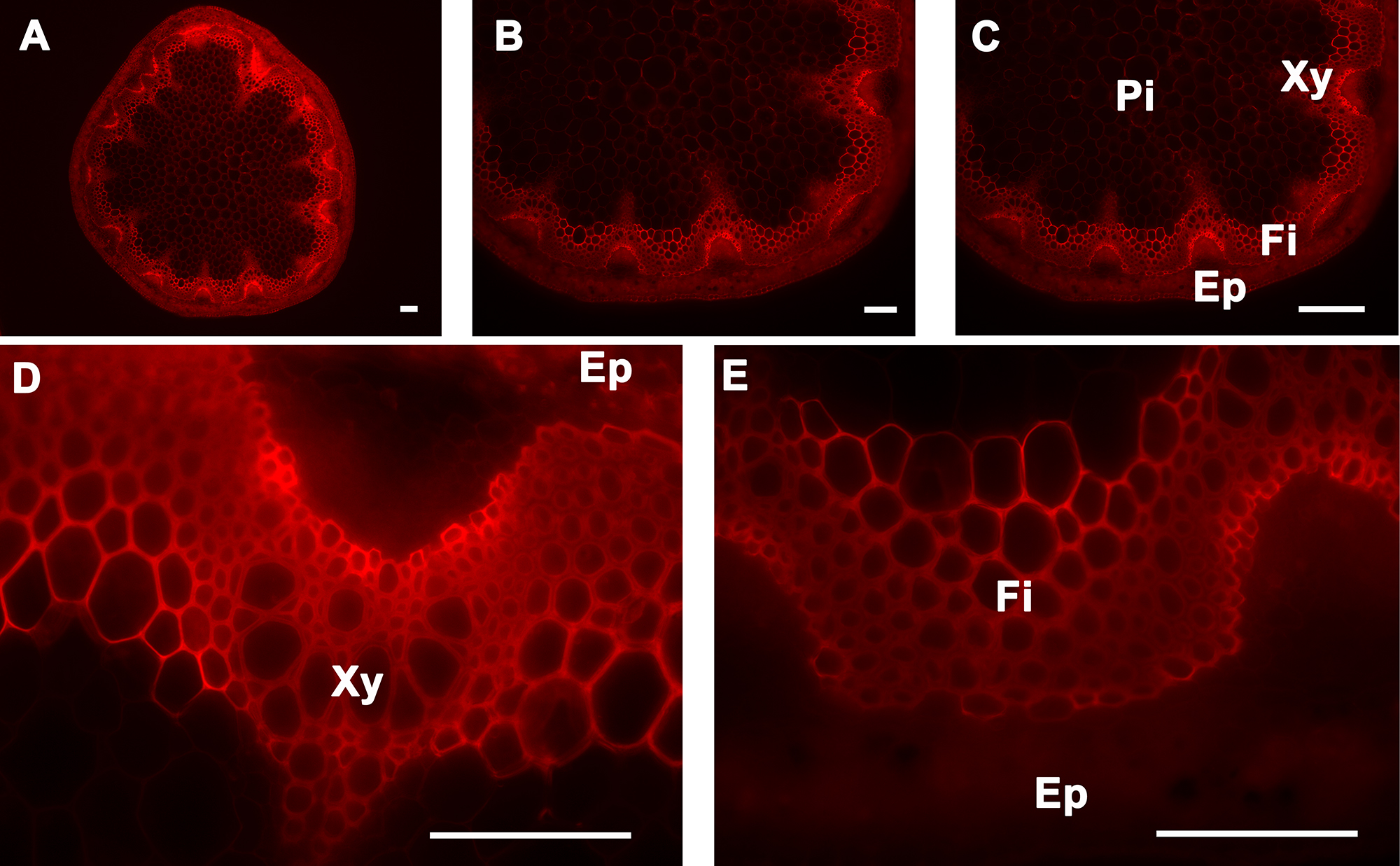
Figure 6. Congo red staining of A. thaliana stem cross-sections. Congo red staining of a wild type A. thaliana stem section showing cellulose deposition in the cell walls of cells of the epidermis, cortex, pith, interfascicular fibers, and xylem (A–E). The positions of the epidermis and cortex (Ep), interfascicular fibers (Fi), pith (Pi), and xylem (Xy) are indicated. Magnifications: Panel A: 5X; Panel B: 10X; Panel C: 20X; Panels D and E: 40X. Bar = 100 µm.
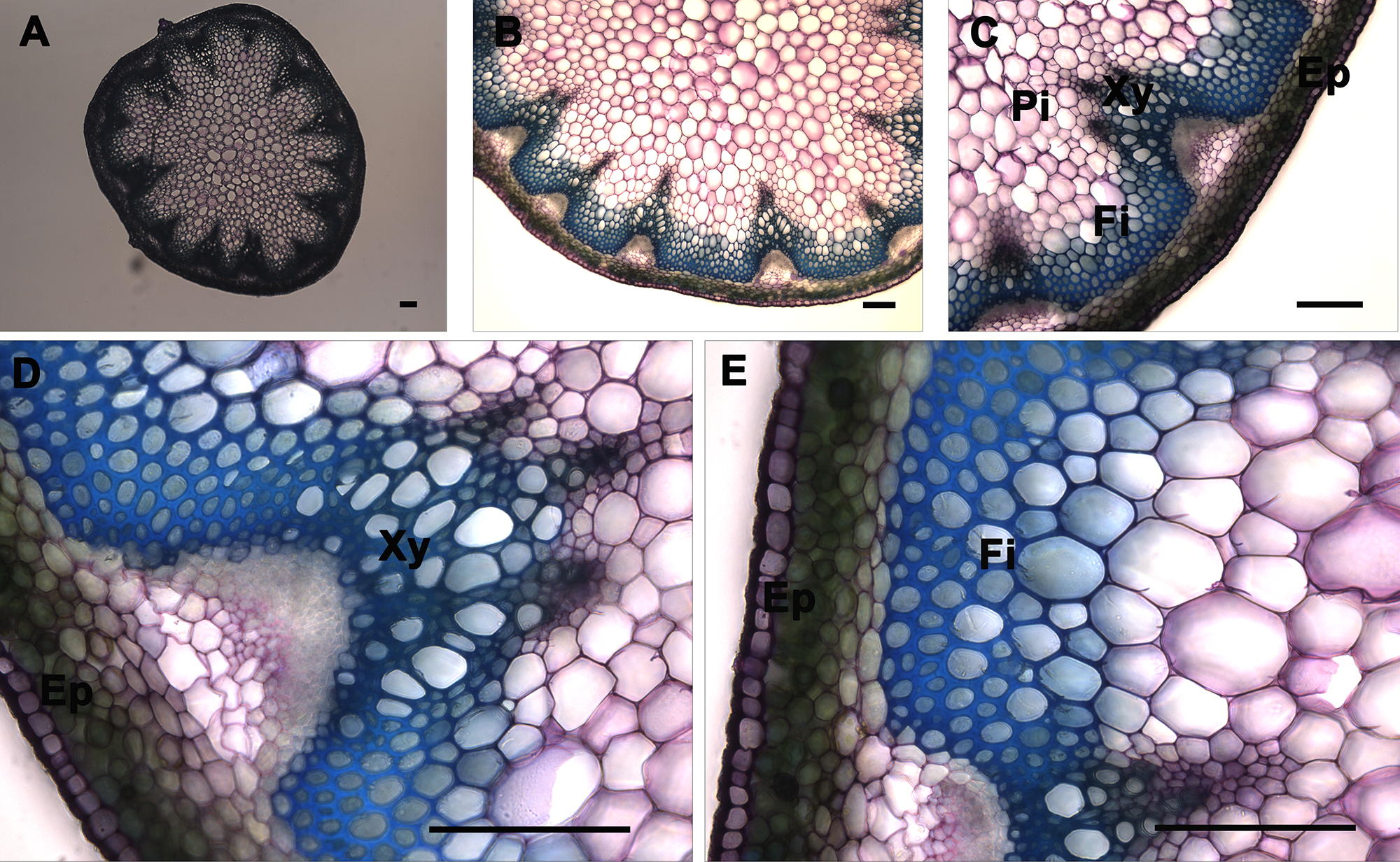
Figure 7. Toluidine blue O staining of A. thaliana stem cross-sections. Toluidine blue O staining of a wild type A. thaliana stem section showing the normal lignin deposition in the walls of interfascicular fibers and xylem cells (A–E). The positions of the epidermis and cortex (Ep), interfascicular fibers (Fi), pith (Pi), and xylem (Xy) are indicated. Magnifications: Panel A: 5X; Panel B: 10X; Panel C: 20X; Panels D and E: 40X. Bar = 100 µm.

Table 1. Microscope and camera settings for imaging. Guidelines for working with fluorescence filters and bright-field imaging. Please click here to view a larger version of this figure.
