Discesa costante La prima indagine mostra alcuna differenza distinguibile nei tassi discendenti della C. elegans durante SWSI usando 633 nm. I tassi discendente sono risultati essere costante a 1,5 mm / sec ± 0.1 mm / sec sia per il vivo e il morto C. elegans. Un campione di 50 worm ha generato una variazione ragionevole del 7% sia per vivere e vermi morti. Non c'è accelerazione agendo sulle viti senza fine poiché la velocità di discesa è costante, in modo che la forza di resistenza è uguale alla forza gravitazionale meno la forza di galleggiamento. Ciò implica che la densità del verme è leggermente più grande di quella dell'acqua; tuttavia, per le stime sottostanti è ancora pratico supporre che la densità di un nematode è all'incirca quella dell'acqua. Modifica della Direzione La seconda indagine, un caso di studio, dimostra che i nematodi sono in grado di cambiare direzione e possono nuotare verso l'alto controgravità. Due curve sono stati montati allo spostamento verticale del nematode modo che le seconde derivate di quelle curve potrebbero essere calettato grafico dell'accelerazione in funzione del tempo (Figura 5). È consigliabile mantenere l'ordine polinomiale più basso possibile, pur mantenendo una buona misura. Un polinomio inferiore indica meno variazione dell'accelerazione nel tempo. I termini polinomiali di ordine superiore saranno trascurabili, e quindi inutile, se l'ordine del polinomio è troppo alta. Questo verme decelera a una velocità costante di 0,110 millimetri / sec 2 ± 0,002 millimetri / sec 2, gira e accelera allo stesso tasso 0,110 millimetri / sec 2 ± 0,002 millimetri / sec 2 fino a poco dopo il punto di svolta. La C. elegans continua a muoversi verso l'alto con un'accelerazione diminuzione di 1,252-,00708 t in mm / sec 2 finché l'accelerazione scende a zero. Tenendo presente Eq. 3, ed una massa verme stimato 3 e #956, g, il worm viene sottoposto ad una forza netta verticale, F Ny, di 0,33 pN fino a poco dopo il punto di svolta. Discesa: Ci sono tre tipi di forze che agiscono sul verme fino a quando il verme raggiunge il punto di svolta: la gravità, trascinare, galleggiabilità e di spinta (Figura 6a). La forza netta è uguale alla somma vettoriale delle tre forze. Qui consideriamo solo i componenti verticali: F Ny = F + F Dy Ty-F g + F B (4) dove F B è la forza di galleggiamento. F g è uguale in grandezza ma opposto in direzione della forza di galleggiamento supponendo che la densità del verme è quello dell'acqua. Eq. 4 può essere scritta nel seguente modo: F Ny = F + F Dy Ty (5) F Ny rimane costante durante la discesa. Ciò implica cheF Dy + F Ty rimane costante fino a quando il nematode raggiunge il punto di svolta. F Dy è più grande nella parte superiore, che è l'inizio del percorso, e si riduce gradualmente fino a zero la velocità è zero al punto svolta mentre F Ty deve aumentare per mantenere costante F Ny. Turnaround: Non c'è forza di trascinamento in senso verticale alla svolta in quanto la velocità verticale pari a zero in quel punto. Le sole forze che agiscono nella direzione verticale sono gravità, – F g, galleggiabilità, F B e il verme spinta verticale, F Ty, come illustrato nella Figura 6b. A questo punto, la spinta della C. elegans possono essere determinate: F Ty = F Ny (6) La spinta in basso della traiettoria è quindi pari a circa 0,33 pN, che è circa 0,001% del peso del verme. Tenendoconto che il peso stimato, F g di C. elegans è 28 nn, Salita: Allo stesso modo, durante la salita, trascinare aumenta ma è puntato verso il basso (Figura 6c): F Ny =-F Dy + F Ty (7) La spinta implementato dal verme deve ora essere ancora più grande e uguale alla somma della forza di resistenza e il peso. Il worm sta rallentando dopo aver iniziato a nuotare in su. Per risalire alla stessa velocità come il verme discese, il verme dovrebbe esercitare una spinta verso l'alto pari ad almeno due volte il suo peso. Librarsi Un esempio di un verme che rallenta la discesa per circa 3 sec è presentato in Figura 7. Un polinomio di 3 ° grado è una misura ragionevole per il percorso complessivo. Gli errori in accelerazione e velocità di questa misura sono inferiori al 15%. Il verme inizia con un significativo upward di spinta, rallenta e comincia a girare intorno a 68 sec; tuttavia, l'accelerazione diminuisce continuamente verso l'alto (Figura 8) fino a quando l'accelerazione netta uguale a zero intorno 68,5 sec. Questo conduce infine a una netta accelerazione nella direzione verso il basso seguito da un altro punto della velocità zero (Figura 9) e il nematode inizia a scendere nuovamente a 69 sec. È interessante notare che l'accelerazione verticale massima osservata in questo caso è di 2,7 mm / sec 2. L'accelerazione nei punti di svolta sono 0,455 millimetri / sec 2 e – 0,455 millimetri / sec, rispettivamente, 2; circa quattro volte più grande che nel caso del nematode che gira e nuota. Uso Eq. 6, si può stimare che la spinta verso l'alto è di circa 1,32 pN al primo punto di svolta. Al secondo punto di svolta, l'accelerazione netto è negativo in modo che la spinta verso l'alto è 1.32 pN. <p class="jove_content" f o: keep-together.within-page = "always"> Figura 1. Impostazione sperimentale. Il laser, il fascio di espansione, l'obiettivo e lo schermo sono essenziali per il setup sperimentale. Gli specchi di sterzo e fori possono essere omessi, ma farà l'allineamento ottico meno stabile. Figura 2. Lunghezza d'onda singola ombra Image (SWSI). Usando 2 mW di luce laser 543 nm l'ombra di un worm viene proiettata su uno schermo. L'immagine viene invertito in modo che il nematode sembra rientrare verso l'alto. 424/51424fig3highres.jpg "src =" / files/ftp_upload/51424/51424fig3.jpg "/> Figura 3. Discesa verticale di un unico tipo selvatico C. elegans in una colonna d'acqua. Questo nematode è stato pedinato da 633 nm luce coerente. La pendenza della regressione lineare indica una velocità di discesa di 1,09 mm / sec ± 0.01 mm / sec. Cliccare qui per vedere una versione più grande di questa figura. Figura 4. Grafico spostamento di un singolo rialzo nuoto di tipo selvatico C. elegans. due attacchi scanalati tracciano il percorso complessivo del nematode. La derivata seconda della forma rivela l'accelerazione. L'accelerazione massima è facilmente desumibile da primo fit: 0,110 millimetri / Sec 2 ± 0,002 millimetri / sec 2. Solo poche barre di errore vengono visualizzati in modo che i punti dati e la vestibilità rimangono visibili. Questo nematode è stato pedinato da 633 nm luce coerente. Cliccare qui per vedere una versione più grande di questa figura. Grafico Figura 5 Accelerazione. Di un unico tipo selvatico C. elegans. Questo nematode mantiene una costante accelerazione verso l'alto di 0,110 millimetri / sec 2 ± 0,002 millimetri / sec 2 inducendolo a rallentare e quindi spostare verso l'alto. Poco dopo il punto di svolta l'accelerazione diminuisce costantemente e l'accelerazione netto diminuisce a zero. g "target =" _blank "> Clicca qui per vedere una versione più grande di questa figura. Diagrammi di Figura 6. Forza per discesa, punto e risalita. Galleggiabilità e la gravità di tornitura sono uguali in modulo e direzione opposta in modo che l'effetto di tali forze si annullano a vicenda. Essi non sono quindi mostrati in questi diagrammi. (A) Il worm sta scendendo con il drag e la spinta verso l'alto. (B) Il worm è al punto più basso della traiettoria senza resistenza. Spinta è rivolta verso l'alto. (C) Il worm è crescente con il drag rivolto verso il basso mentre la spinta è rivolta verso l'alto. "Src =" / files/ftp_upload/51424/51424fig7.jpg "/> Figura 7. Grafico spostamento del singolo nematode bilico. Il worm rallenta e comincia a girare intorno a circa 68 secondi, ma inizia a scendere intorno al 69 sec. Cliccare qui per vedere una versione più grande di questa figura. .. Figura 8 grafico accelerazione di un singolo nematode bilico Questo worm inizia con un'accelerazione verso l'alto di 2,7 millimetri / sec 2, riduce a zero e, infine, ha una accelerazione verso il basso di -. 2,6 millimetri / sec 2 Cliccare qui per vedere un grande versione di questa figura. Figura 9. Grafico della velocità di un singolo nematode bilico. Questo worm si ferma a 68 sec e inizia a dirigersi verso l'alto, rallenta e raggiunge la velocità zero nella direzione verticale a 69 sec seguita da una discesa. Cliccare qui per visualizzare un grande versione di questa figura. Tabella 1. Velocità media di discesa di N2 C. elegans. vivo 50 e 50 nematodi morti sono tracked durante la loro discesa. La velocità media per le discese è lo stesso per la vita e le vermi morti: 1,5 mm / sec ± 0.1 mm / sec.
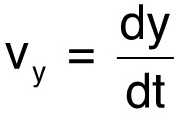 (1)
(1) 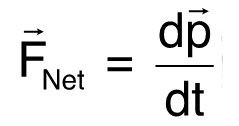 (2)
(2) 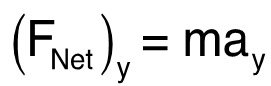 (3)
(3) 