किसी भी जीवित जीव के जीनोम इंजीनियर करने की क्षमता कई जैव चिकित्सा और जैव प्रौद्योगिकीय अनुप्रयोगों, जैसे रोग के सुधार के रूप में परिवर्तन, रोग अध्ययन के लिए सटीक सेलुलर मॉडल के निर्माण, या कृषि के उत्पादन वांछित लक्षण के साथ फसलों । सदी की बारी के बाद से, स्तनधारी कोशिकाओं में जीनोम इंजीनियरिंग के लिए विभिन्न प्रौद्योगिकियों का विकास किया गया है, जिनमें मेगान्यूक्लीसिस1,2,3, जिंक फिंगर न्यूक्लिसेस4,5, या प्रतिलेखन उत्प्रेरक-जैसे अमोघ नाभिकों (टैलेंस)6,7,8,9. हालांकि, इन पहले प्रौद्योगिकियों या तो कार्यक्रम के लिए मुश्किल है या इकट्ठा करना कठिन है, जिससे अनुसंधान और उद्योग में अपने व्यापक अपनाने बाधा ।
हाल के वर्षों में, संकुल नियमित रूप से interspaced लघु palindromic दोहराता (CRISPR)-CRISPR-एसोसिएटेड (कैस) प्रणाली एक शक्तिशाली नए जीनोम इंजीनियरिंग प्रौद्योगिकी10,11के रूप में उभरा है । मूल रूप से बैक्टीरिया में एक अनुकूली प्रतिरक्षा प्रणाली, यह सफलतापूर्वक पौधों और जानवरों, मनुष्यों सहित में जीनोम संशोधन के लिए तैनात किया गया है । एक प्राथमिक कारण CRISPR-कैस इतने कम समय में इतनी लोकप्रियता प्राप्त की है कि तत्व है कि Cas9 या Cas12a (भी Cpf1 के रूप में जाना जाता है) के रूप में महत्वपूर्ण कैस endonuclease, लाता है, जीनोम में सही स्थान के लिए बस चिमेरिक एकल गाइड का एक छोटा टुकड़ा है RN एक (sgRNA), जो डिजाइन और सस्ते synthesize करने के लिए सीधा है । लक्ष्य स्थल पर भर्ती होने के बाद, कैस एंजाइम आणविक कैंची और cleaves की एक जोड़ी के रूप में कार्य करता है अपने ruvc, hnh, या nuc डोमेन12,13,14के साथ बंधे डीएनए । परिणामस्वरूप डबल फंसे तोड़ (DSB) बाद में या तो गैर-समजातीय अंत में शामिल होने (NHEJ) या homology-निर्देशित मरंमत (HDR) मार्ग के माध्यम से कोशिकाओं द्वारा मरंमत की है । एक मरंमत टेम्पलेट के अभाव में, DSB त्रुटि-प्रवण NHEJ मार्ग द्वारा सुधारा गया है, जो कटौती साइट पर छद्म यादृच्छिक प्रविष्टि या न्यूक्लियोटिडों (indels) के विलोपन को जंम दे सकता है, संभवतः प्रोटीन-कोडिंग जीन में frameshift म्यूटेशन के कारण । हालांकि, एक दाता टेंपलेट है कि वांछित डीएनए परिवर्तन शामिल की उपस्थिति में, DSB उच्च निष्ठा HDR मार्ग द्वारा मरंमत की है । दाता टेम्पलेट्स के आम प्रकार एकल-कतरा हुआ oligonuक्लिओटाइड्स (ssODNs) और plasmids शामिल हैं । पूर्व आम तौर पर अगर इरादा डीएनए परिवर्तन (उदाहरण के लिए, एक एकल आधार जोड़ी के परिवर्तन) छोटे है प्रयोग किया जाता है, जबकि बाद आम तौर पर अगर एक एक अपेक्षाकृत लंबे अनुक्रम (उदाहरण के लिए, एक हरी फ्लोरोसेंट प्रोटीन के कोडन अनुक्रम डालने की इच्छा है प्रयोग किया जाता है जीएफपी) लक्ष्य लोकस में ।
कैस प्रोटीन की एंडोन्यूक्लिएज गतिविधि के लिए लक्ष्य स्थल15पर एक प्रोटोस्पेसर सटी आकृति (पाम) की उपस्थिति की आवश्यकता होती है । Cas9 का पाम protospacer के 3 ‘ अंत में है, जबकि Cas12a के पाम (भी Cpf1 कहा जाता है) 5 ‘ अंत में16के बजाय है । कैस-गाइड आरएनए परिसर में एक DSB पेश करने में असमर्थ है अगर पाम अनुपस्थित है17। अतः पाम उन जीनोमिक स्थानों पर एक बाधा रखता है जहाँ एक विशेष कैस नाभिक सट कर आ जाता है । सौभाग्य से, विभिन्न जीवाणु प्रजातियों से कैस नाभिक आमतौर पर विभिन्न पाम आवश्यकताओं का प्रदर्शन. इसलिए, हमारे इंजीनियरिंग toolbox में विभिन्न CRISPR-Cas सिस्टम को एकीकृत करके, हम एक जीनोम में लक्षित किया जा सकता है कि साइटों की सीमा का विस्तार कर सकते हैं । इसके अलावा, एक प्राकृतिक कैस एंजाइम इंजीनियर या वैकल्पिक पाम दृश्यों को पहचानने के लिए विकसित किया जा सकता है, और अधिक हेरफेर करने के लिए सुलभ जीनोमिक लक्ष्यों की गुंजाइश को चौड़ा करने18,19,20।
हालांकि कई CRISPR-कैस सिस्टम जीनोम इंजीनियरिंग प्रयोजनों के लिए उपलब्ध हैं, प्रौद्योगिकी के अधिकांश उपयोगकर्ताओं को Cas9 nuclease पर मुख्य रूप से कई कारणों के लिए स्ट्रेप्टोकोकस pyogenes (SpCas9) से भरोसा किया है । सबसे पहले, यह एक अपेक्षाकृत बस NGG पाम की आवश्यकता है, कई अंय कैस प्रोटीन के विपरीत है कि केवल अधिक जटिल PAMs की उपस्थिति में सट कर सकते हैं । दूसरा, यह मानव कोशिकाओं21,22,23,24में सफलतापूर्वक तैनात किया जाने वाला पहला कैस एंडोन्यूक्लिएज है । तीसरा, SpCas9 अब तक की तारीख को सबसे अच्छी विशेषता एंजाइम है । यदि एक शोधकर्ता एक और कैस nuclease का उपयोग करना चाहता है, वह या वह अक्सर कैसे सबसे अच्छा प्रयोग डिजाइन और कितनी अच्छी तरह अंय एंजाइमों अलग जैविक संदर्भों में SpCas9 की तुलना में प्रदर्शन करेंगे के बारे में स्पष्ट नहीं होगा ।
अलग CRISPR-Cas सिस्टम के सापेक्ष प्रदर्शन के लिए स्पष्टता प्रदान करने के लिए, हम हाल ही में पांच Cas endonucleases की एक व्यवस्थित तुलना प्रदर्शन किया है – SpCas9, Staphylococcus औरेअस से Cas9 एंजाइम (SaCas9), Cas9 एंजाइम से नेइससेरिया मेनिनगिटिडिस (NmCas9), एसिडमिनोकोकस एसपी. BV3L6 (AsCas12a) से Cas12a एंजाइम, और लच्नोस्पाइरेसी बैक्टीरियम Cas12a (ND2006)25से LbCas12a एंजाइम । एक निष्पक्ष तुलना के लिए, हम विभिंन कैस नाभिक लक्ष्य साइटों और अंय प्रायोगिक शर्तों के एक ही सेट का उपयोग कर मूल्यांकन किया । अध्ययन में प्रत्येक CRISPR-Cas प्रणाली के लिए डिजाइन पैरामीटरों को भी चित्रित किया गया है, जो प्रौद्योगिकी के प्रयोक्ताओं के लिए उपयोगी संदर्भ के रूप में कार्य करेगा । यहां, बेहतर शोधकर्ताओं CRISPR-कैस प्रणाली का उपयोग करने के लिए सक्षम करने के लिए, हम अलग Cas9 और Cas12a एंजाइमों के साथ इष्टतम जीनोम इंजीनियरिंग के लिए एक कदम दर कदम प्रोटोकॉल प्रदान ( चित्रा 1देखें) । प्रोटोकॉल न केवल प्रयोगात्मक विवरण पर भी महत्वपूर्ण डिजाइन विचार शामिल स्तनधारी कोशिकाओं में एक सफल जीनोम इंजीनियरिंग परिणाम की संभावना को अधिकतम करने के लिए ।
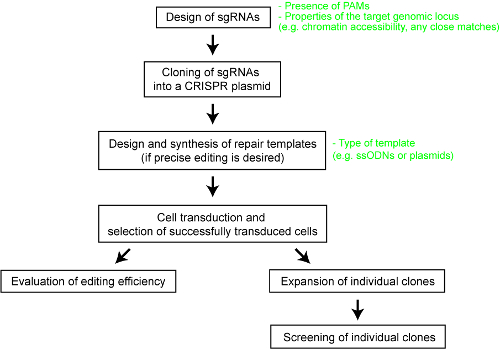
चित्रा 1 : जीनोम संपादित मानव कोशिका रेखाएँ उत्पन्न करने के लिए कार्यप्रवाह का अवलोकन. कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें ।
