Способность инженер геном любого живого организма имеет много биомедицинских и биотехнологических приложений, например коррекции болезнетворные мутации, строительство точных моделей для исследования заболеваний, или поколения сельскохозяйственных культур с желательные черты. С начала века, различные технологии были разработаны для техники генома клеток млекопитающих, включая meganucleases1,2,3, цинковый палец nucleases4,5, или Транскрипция эффекторных активатор как nucleases (Таленс)6,,78,9. Однако эти более ранних технологий трудно программы или утомительно собрать, препятствуя тем самым их широкое распространение в научных исследований и промышленности.
В последние годы кластеризованный регулярно interspaced короткие палиндром повторяется (ТРИФОСФАТЫ) – связанные ТРИФОСФАТЫ (Cas) системы стала мощной новой генома, инженерные технологии10,11. Первоначально адаптивной иммунной системы в бактерии, он был успешно развернут для модификации генома растений и животных, включая человека. Основная причина, почему ТРИФОСФАТЫ-Cas популярность так много за такое короткое время, что элемент, который приносит ключевых эндонуклеазы Cas, например Cas9 или Cas12a (также известный как Cpf1), в нужное место в геноме это просто небольшой фрагмент химерных единого руководства RN A (sgRNA), которая является простой дизайн и дешевые синтезировать. После набираются на целевой сайт, Cas фермента функционирует как молекулярные ножницы и расщепляет ДНК связанного с его RuvC, HNH или КНУ доменов12,,1314. Результате двойной мель перерыв (DSB) впоследствии отремонтированы клетки через ориентированные на гомологии ремонт (HDR) путь или не гомологичных конце присоединения (NHEJ). В отсутствие шаблона ремонт DSB отремонтированы по ошибкам NHEJ пути, который может привести к псевдо-случайных вставки или удаления нуклеотидов (indels) в месте разреза, потенциально вызывая фреймшифт мутации в генах, белок кодирования. Однако в присутствии доноров шаблон, содержащий требуемые изменения ДНК, DSB восстановлена дорожка HDR высокой верности. Общие типы доноров шаблоны включают одноцепочечной олигонуклеотиды (ssODNs) и плазмид. Бывший обычно используется, если предполагаемого изменения ДНК небольшие (например, изменение одной пары базы), в то время как последние обычно используется, если один хочет, чтобы вставить относительно длинная последовательность (например, кодирующая последовательность зеленого флуоресцентного белка или GFP) в целевой Локус.
Эндонуклеазы активность белка Cas требует наличия protospacer прилегающие мотив (PAM) на целевой сайт15. Пэм Cas9 находится в конце 3′ protospacer, в то время как Пэм Cas12a (также называемый Cpf1) находится в конце 5′ вместо16. Cas руководство РНК комплекс не в состоянии представить DSB, если PAM отсутствует17. Следовательно PAM места ограничения в геномной местах, где особенно нуклеиназы Cas способен расщеплять. К счастью Cas nucleases из различных видов бактерий обычно демонстрируют различные требования PAM. Таким образом путем интеграции различных систем ТРИФОСФАТЫ-Cas в наших инженерных инструментов, мы можем расширить круг сайтов, которые могут быть направлены в геноме. Кроме того естественный фермент Cas может быть спроектирован или эволюционировали признать альтернативных последовательностей PAM, дальнейшего расширения масштабов геномной целей доступным для манипуляции18,19,20.
Хотя несколько ТРИФОСФАТЫ-Cas системы доступны для технических целей генома, большинство пользователей технологии полагались главным образом на Cas9 Нуклеаза от Streptococcus pyogenes (SpCas9) по нескольким причинам. Во-первых он требует относительно просто NGG Пэм, в отличие от многих других белков Cas, которые может только расщеплять при наличии более сложных пам. Во-вторых это первый Cas эндонуклеазы успешно развернуты в клетки человека21,,2223,24. В-третьих SpCas9 является на сегодняшний день лучше всего характеризуется фермент на сегодняшний день. Если исследователь хочет использовать другой нуклеиназы Cas, он или она часто будет ясно о том, как лучше разработать эксперимент, и насколько хорошо другие ферменты будет выполнять в различных биологических условиях по сравнению с SpCas9.
Чтобы внести ясность в относительной эффективности различных систем ТРИФОСФАТЫ-Cas, мы провели недавно систематическое сравнение пяти Cas эндонуклеазами – SpCas9, Cas9 фермента от золотистого стафилококка (SaCas9), энзим Cas9 от Neisseria meningitidis (NmCas9), Cas12a фермента от Acidaminococcus sp. BV3L6 (AsCas12a) и Cas12a фермента от бактерия Lachnospiraceae ND2006 (LbCas12a)25. За справедливое сравнение мы оценивали различные Cas nucleases, используя тот же набор целевых объектов и других экспериментальных условий. Исследование также определила дизайн параметры для каждой системы ТРИФОСФАТЫ-Cas, которая будет служить полезным справочным материалом для пользователей технологии. Здесь, чтобы лучше включить исследователей использовать ТРИФОСФАТЫ-Cas системы, мы предоставляем пошаговые протокол для оптимального генома техники с различными Cas9 и Cas12a ферменты (см. Рисунок 1). Протокол включает не только экспериментальных детали, но также важных конструктивных соображений необходимо максимизировать вероятность успешного генома инженерных решений в mammalian клетках.
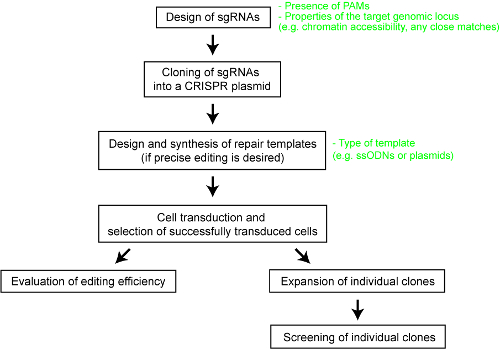
Рисунок 1 : Обзор рабочего процесса для создания генома линий клеток человека отредактированы. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
