القدرة على هندسة الجينوم من أي كائن حي لديه العديد من التطبيقات الطبية الحيوية والتكنولوجيا الحيوية، مثل تصحيح الأمراض المسببة للطفرات، وبناء النماذج الخلوية الدقيقة لدراسات الأمراض، أو توليد الزراعية المحاصيل مع الصفات المرغوب فيه. منذ مطلع هذا القرن، تم استحداث تكنولوجيات مختلفة لهندسة الجينوم في خلايا الثدييات، بما في ذلك ميجانوكليسيس1،2،3، زنك إصبع نوكليسيس4،5، أو النسخ المستجيب المنشط–مثل نوكليسيس (تالينس)6،7،،من89. هذه التكنولوجيات السابقة غير صعبة للبرنامج أو شاقة التجمع، مما يعوق اعتمادها على نطاق واسع في البحوث والصناعة.
في السنوات الأخيرة، بشكل منتظم مجمع إينتيرسباسيد في يكرر المتناوب قصيرة (كريسبر)-برز نظام المرتبطة كريسبر (Cas) جينوم جديدة قوية هندسة تكنولوجيا10،11. أصلاً نظام المناعة تكيفية في البكتيريا، قد تم بنجاح نشر لتعديل الجينوم في النباتات والحيوانات، بما فيها البشر. سبب الرئيسي لماذا كريسبر Cas قد اكتسب شعبية الكثير في وقت قصير كهذا هو أن العنصر الذي يجلب endonuclease الأكاديمية الرئيسية، مثل Cas9 أو Cas12a (المعروفة أيضا باسم Cpf1)، إلى الموقع الصحيح في الجينوم هو مجرد قطعة قصيرة من دليل واحد تشيميريك RN (سجرنا)، وهي واضحة للتصميم ورخيصة توليف. بعد تجنيدهم إلى الموقع المستهدف، تعمل مقص الجزيئية الإنزيم Cas وكليافيس الحمض النووي المرتبطة مع به روفك، HNH، أو نشاط المجالات12،،من1314. بعد ذلك يتم إصلاح كسر الذين تقطعت بهم السبل مزدوجة الناتجة (جهاز تسوية المنازعات) بالخلايا عن طريق نهاية غير المتجانسة الالتحاق بالعمل (نج) أو مسار الإصلاح الموجه من التماثل (HDR). في غياب إصلاح قالب، يتم إصلاح جهاز تسوية المنازعات بالمسار نهيج عرضه للخطأ، ويمكن أن يؤدي إلى شبه عشوائي الإدراج أو الحذف من النيوكليوتيدات (إينديلس) في موقع قص، يحتمل أن تسبب فراميشيفت الطفرات في الجينات ترميز البروتين. ومع ذلك، حضور الجهات مانحة قالب يحتوي على الحمض النووي التغييرات المطلوبة، يتم إصلاح جهاز تسوية المنازعات بالمسار HDR عالية الدقة. وتشمل الأنواع الشائعة من قوالب المانحة النوكليوتيد المفرد-الذين تقطعت بهم السبل (سودنس) ووالبلازميدات. عادة ما تستخدم في السابق إذا كانت التغييرات الحمض النووي المقصود صغيرة (على سبيل المثال، تغيير زوج قاعدة واحدة)، في حين أن هذا الأخير يستخدم عادة إذا كان أحد يرغب في إدراج تسلسل طويلة نسبيا (على سبيل المثال، تسلسل ترميز بروتين فلوري أخضر أو التجارة والنقل) إلى المكان المستهدف.
النشاط اندونوكليسي من البروتين Cas يتطلب وجود بروتوسباسير عزر المتاخمة (بام) في الموقع المستهدف15. بام Cas9 في نهاية 3 ‘بروتوسباسير، بينما بام Cas12a (وتسمى أيضا Cpf1) في النهاية 5’ بدلاً من16. Cas-دليل الحمض النووي الريبي المعقدة غير قادر على عرض جهاز تسوية المنازعات إذا تغيب17بام. ومن ثم، بام يضع قيداً على مواقع الجينوم قادرة على تنشق فيه نوكلاس Cas خاصة. لحسن الحظ، يحمل nucleases الأكاديمية الصينية للعلوم من مختلف الأنواع البكتيرية عادة متطلبات مختلفة بام. ومن ثم، بإدماج مختلف Cas كريسبر نظم لدينا الأدوات الهندسية، نحن توسيع نطاق المواقع التي قد تكون مستهدفة في جينوم. وعلاوة على ذلك، يمكن إجراء هندسة عكسية إنزيم الأكاديمية الصينية للعلوم طبيعية أو تطورت إلى الاعتراف بتسلسل بام بديلة، مزيد من توسيع نطاق أهداف الجينوم موجوداً للتلاعب18،،من1920.
على الرغم من أن تتوفر أنظمة كريسبر Cas متعددة الأغراض الهندسية الجينوم، اعتمدت معظم المستخدمين لهذه التكنولوجيا أساسا على نوكلاس Cas9 من العقدية المقيّحة (SpCas9) لأسباب متعددة. أولاً، فإنه يتطلب أم نج نسبيا ببساطة، خلافا للعديد من البروتينات Cas الأخرى التي يمكن أن تهزم سوى حضور PAMs أكثر تعقيداً. وثانيا، أنها endonuclease الأكاديمية الأولى التي سيتم نشرها بنجاح في الخلايا البشرية21،22،،من2324. وثالثاً، SpCas9 هو إلى حد بعيد أفضل الإنزيم تتسم حتى الآن. إذا لم يرغب باحث في استخدام آخر Cas نوكلاس، أنه أو أنها غالباً ما ستكون واضحة بشأن أفضل السبل لتصميم التجربة وجيدا كيف سوف تؤدي الإنزيمات الأخرى في مختلف السياقات البيولوجي مقارنة SpCas9.
توفير الوضوح لأداء مختلف Cas كريسبر النظم النسبية، قد أجرينا مؤخرا مقارنة منهجية لخمسة Cas اندونوكليسيس-SpCas9، الإنزيم Cas9 من المكوّرات العنقودية الذهبية (SaCas9)، الإنزيم Cas9 من وجود النيسرية السحائية (NmCas9)، الإنزيم Cas12a من أسيدامينوكوككوس sp. BV3L6 (AsCas12a)، والانزيم Cas12a من بكتيريا لاتشنوسبيراسيي ND2006 (LbCas12a)25. لمقارنة منصفة، يمكننا تقييم nucleases Cas مختلفة باستخدام نفس مجموعة من المواقع المستهدفة وغيرها من الشروط التجريبية. الدراسة أيضا تحديد معايير التصميم لكل نظام كريسبر-الأكاديمية الصينية للعلوم، الذي سيكون بمثابة مرجع مفيد للمستخدمين للتكنولوجيا. هنا، على نحو أفضل تمكن الباحثين من الاستفادة من كريسبر-الأكاديمية الصينية للعلوم نظام، نحن نقدم بروتوكول خطوة بخطوة لهندسة الجينوم الأمثل مع مختلف الإنزيمات Cas9 و Cas12a (انظر الشكل 1). ليس فقط يتضمن البروتوكول التجريبي من التفاصيل لكن الاعتبارات الخاصة بالتصميم أيضا أقصى حد احتمال نتيجة هندسة الجينوم ناجحة في خلايا الثدييات.
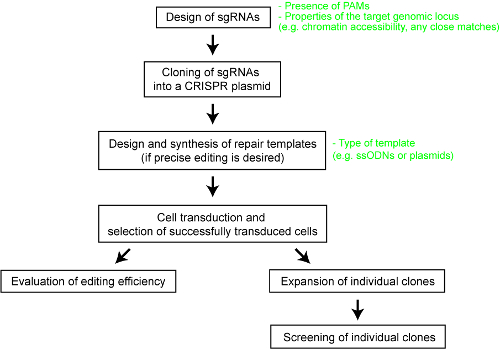
الشكل 1 : لمحة عامة عن سير العمل لتوليد الجينوم تحرير خطوط الخلايا البشرية- الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
