La capacité à l’ingénieur le génome de tout organisme vivant a beaucoup d’applications biomédicales et biotechnologiques, telles que la correction des pathogènes mutations, construction de modèles cellulaires précis pour les études de la maladie, ou génération d’agricoles cultures avec traits désirables. Depuis le tournant du siècle, plusieurs technologies ont été développées pour le génie du génome dans les cellules de mammifères, y compris les méganucléases1,2,3, zinc finger nucleases4,5, ou transcription activator comme effecteur nucléases (TAPS)6,7,8,9. Cependant, ces technologies antérieures sont difficiles à programmer ou fastidieux à monter, ce qui entrave leur adoption généralisée dans la recherche et l’industrie.
Ces dernières années, le cluster régulièrement ponctuées de courtes répétitions palindromiques (CRISPR) – système associées à CRISPR (Cas) a émergé comme un puissant génome nouvel ingénierie technologie10,11. Initialement un système immunitaire adaptatif chez les bactéries, il a été correctement déployé pour la modification du génome chez les plantes et les animaux, y compris les humains. Des principales raisons pourquoi CRISPR-Cas a gagné en popularité autant en si peu de temps qui est l’élément qui apporte l’endonucléase de Cas clé, tels que Cas9 ou Cas12a (également connu comme Cpf1), à l’emplacement correct dans le génome est simplement un petit morceau de guide unique chimérique RN A (sgRNA), qui est simple de conception et pas cher à synthétiser. Après recrutement vers le site cible, l’enzyme Cas fonctionne comme une paire de ciseaux moléculaires et fend l’ADN lié avec son RuvC, HNH ou Nuc domaines12,13,14. La rupture de brin double (DSB) qui en résulte est réparée par la suite par les cellules via la fin non homologue rejoindre (NHEJ) ou voie de réparation axés sur l’homologie (HDR). En l’absence d’un modèle de la réparation, l’ORD est réparé par la voie NHEJ erreurs, qui peut donner lieu à pseudo-aléatoire d’insertion ou de suppression de nucléotides (indels) sur le site de la coupe, causant potentiellement déphasage des mutations dans les gènes codant pour des protéines. Toutefois, en présence d’un modèle de donateurs qui contient les modifications souhaitées de l’ADN, l’ORD est réparé par la voie du HDR haute fidélité. Types communs de modèles de donateurs comprennent des oligonucléotides monocaténaires (ssODNs) et plasmides. Le premier est généralement utilisé si les changements prévus de l’ADN sont de petite taille (par exemple, modification d’un seul nucléotide), alors que ce dernier est généralement utilisé si l’on veut introduire une séquence relativement longue (par exemple, la séquence codante d’une protéine fluorescente verte ou GFP) dans le locus de la cible.
L’activité endonucléasique de la protéine Cas nécessite la présence d’un motif adjacent de protospacer (PAM) à la cible site15. Le PAM de Cas9 est à l’extrémité 3′ de la protospacer, tandis que le PAM de Cas12a (également appelé Cpf1) est à l’extrémité 5′ à la place16. Le Cas-guide RNA complexe ne peut pas introduire un ORD si la PAM est absente17. Par conséquent, la PAM place une contrainte sur les génomiques emplacements où une nucléase de Cas particulier est capable de cliver. Heureusement, nucléases Cas de différentes espèces de bactéries présentent généralement des exigences différentes de PAM. Par conséquent, en intégrant différents systèmes CRISPR-Cas dans notre boîte à outils technique, nous pouvons élargir l’éventail des sites qui peuvent être ciblés dans un génome. En outre, une enzyme naturelle de Cas peut être conçue ou évoluée pour reconnaître les alternatives des séquences de PAM, encore élargir la portée de génomiques cibles accessibles aux manipulations18,19,20.
Bien que plusieurs systèmes CRISPR-Cas soient disponibles pour fins d’ingénierie du génome, la plupart des utilisateurs de la technologie sont appuient principalement sur la nucléase Cas9 de Streptococcus pyogenes (SpCas9) pour de multiples raisons. Tout d’abord, il faut un PAM relativement simplement NGG, contrairement à plusieurs autres protéines de Cas qui peuvent s’attacher uniquement en présence de SDS plus complexes. Deuxièmement, il est le premier endonucléase de Cas qui seront déployés avec succès dans les cellules humaines21,22,23,24. En troisième lieu, le SpCas9 est de loin la meilleure enzyme caractérisé à ce jour. Si un chercheur souhaite utiliser la nucléase un autre Cas, il ou elle serait souvent peu claire sur la meilleure façon de concevoir l’expérience et comment bien les autres enzymes seront produira dans différents contextes biologiques par rapport à SpCas9.
Pour fournir la clarté à la performance relative des différents systèmes CRISPR-Cas, nous avons récemment effectué une comparaison systématique des cinq endonucléases de Cas – SpCas9, l’enzyme Cas9 Staphylococcus aureus (SaCas9), l’enzyme Cas9 de Neisseria meningitidis (NmCas9), l’enzyme Cas12a de Acidaminococcus SP. BV3L6 (AsCas12a) et l’enzyme Cas12a de bactérie Lachnospiraceae ND2006 (LbCas12a)25. Pour une comparaison équitable, nous avons évalué les nucléases de Cas différents, en utilisant le même ensemble de sites cibles et autres conditions expérimentales. Les paramètres de conception d’étude aussi délimité pour chaque système de SAE-CRISPR, qui servirait de référence utile pour les utilisateurs de la technologie. Ici, pour mieux permettre aux chercheurs de faire usage de la SAE-CRISPR système, nous fournissons un protocole étape par étape pour le génie de génome optimale avec des enzymes de Cas9 et Cas12a différentes (voir Figure 1). Le protocole inclut non seulement les détails expérimentaux, mais les considérations de conception aussi important pour maximiser la probabilité d’un résultat technique génome réussie dans les cellules de mammifères.
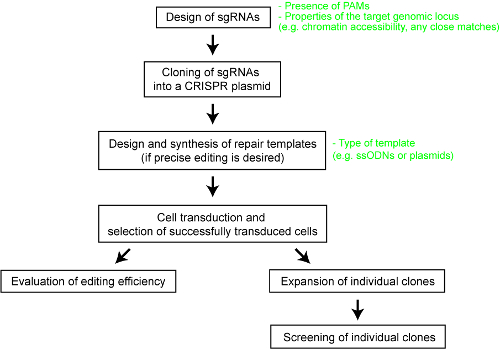
Figure 1 : Une vue d’ensemble du flux de travail pour générer du génome édité des lignées cellulaires humaines. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
