Hastalığa neden olan düzeltilmesi gibi birçok Biyomedikal ve Biyoteknolojik uygulamaları herhangi bir canlı organizma genom mühendis yeteneğine sahiptir mutasyonlar, hastalık çalışmalar için doğru hücresel modelleri inşaatı veya, tarımsal üretimi arzu edilen özellikleri ile bitkileri. Yüzyılın beri genom Mühendisliği dahil meganucleases1,2,3memeli hücrelerinde için çeşitli teknolojiler geliştirilmiştir, parmak nükleaz4,5, çinko veya Transkripsiyon harekete geçirmek gibi efektör enzimler (TALENs)6,7,8,9. Ancak, bu önceki böylece onların yaygınlaşmasının araştırma ve endüstrisi engelleyici programı için zor ya da sıkıcı bir araya getirmek teknolojilerdir.
Son yıllarda, kümelenmiş düzenli olarak kısa palindromik yineler (CRISPR) interspaced – CRISPR-ilişkili (Cas) sistem teknoloji10,11mühendislik güçlü yeni genom ortaya çıkmıştır. Aslında bir adaptif bağışıklık sisteminde bakteri, başarıyla oldu genom değişiklik bitkiler ve hayvanlar, insanlar da dahil olmak üzere dağıtılmış. Neden CRISPR-Cas kısa bir sürede çok popülerlik kazanmıştır bir temel nedeni bu Cas9 veya Cas12a (Ayrıca Cpf1 da bilinir) gibi anahtar Cas endonükleaz getiriyor öğe, genom doğru konuma chimeric tek Kılavuzu RN sadece kısa bir parçası (SgRNA), bir tasarım için basit ve ucuz sentezlemek. Hedef siteye işe ediliyor sonra CA’ları enzim moleküler makas çifti olarak işlev görür ve onun RuvC, HNH veya Nuc etki alanları12,13,14ile ilişkili DNA cleaves. Elde edilen çift telli sonu (DSB) daha sonra homolog olmayan sonu (NHEJ) katılma veya homoloji yönetmen onarım (HDR) yolu ile hücreleri tarafından onarıldı. Onarım şablon yokluğunda, DSB potansiyel olarak protein kodlayıcı genlerin frameshift mutasyonlar neden çıkmasına neden takma ad-rasgele ekleme veya silmek-in nükleotit (indels) kesim yerinde olabilir, hataya NHEJ yolu tarafından onarıldı. Ancak, istediğiniz DNA değişiklikleri içeren bir donör şablonu huzurunda DSB yüksek sadakat HDR yolu tarafından onarıldı. Donör şablonları sık karşılaşılan tek iplikçikli oligonucleotides (ssODNs) ve plazmid bulunmaktadır. Eğer bir nispeten uzun bir sıra eklemek dilek ikinci genellikle kullanılırken dilediğiniz DNA değişiklikleri (örneğin, tek bir baz çifti İLETİMLERİNİZE), küçük eski genellikle kullanılır (örneğin, bir yeşil flüoresan protein kodlama dizisi veya GFP) hedef locus içine.
CA’ları protein endonükleaz Etkinlik hedef site15bir protospacer bitişik motifi (PAM) olmasını gerektirir. (Cpf1 olarak da bilinir) Cas12a PAM 5′ sonunda bunun yerine olmakla Cas9 PAM163′ sonunda protospacer olduğunu. Cas-Kılavuzu RNA karmaşık PAM17ise bir DSB tanıtmak değiştiremiyor. Bu nedenle, PAM belirli bir Cas nükleaz ayırmak mümkün nerede genomik Mekanlar üzerinde bir kısıtlama yerleştirir. Neyse ki, Cas enzimler farklı bakteri türleri üzerinden genellikle farklı PAM gereksinimleri sergi. Bu nedenle, çeşitli CRISPR-Cas sistemleri mühendislik bizim alet entegre ederek, bir genom hedefleyebilir siteleri aralığını genişletebilirsiniz. Ayrıca, doğal bir Cas enzim mühendislik veya daha fazla genomik hedefleri manipülasyon18,19,20‘ ye erişilebilir kapsamını genişletme alternatif PAM dizileri tanımasını gelişti.
Teknolojisi çoğu kullanıcı için birden çok nedeni birden çok CRISPR-Cas sistemi genom mühendislik amaçları için kullanılabilir olmakla birlikte, ağırlıklı olarak Cas9 nükleaz Streptococcus pyogenes (SpCas9) üzerinden yararlanmıştır. Birincisi, bir nispeten sade bir şekilde NGG PAM, aksine sadece varlığında daha karmaşık PAMs bölmek birçok diğer Cas protein gerektirir. İkincisi, bu insan hücreleri21,22,23,24‘ te başarılı bir şekilde dağıtılması için ilk Cas endonükleaz ‘s. Üçüncü olarak, SpCas9 bugüne kadar en iyi karakterize enzim gereğidir. Bir araştırmacı başka bir Cas nükleaz kullanmak isterse, o ya da o sık sık tasarım deneme ve nasıl su kuyusu diğer enzimler için SpCas9 göre farklı biyolojik bağlamlarda yapmak için en iyi nasıl hakkında belirsiz olur.
Farklı CRISPR-Cas sistemleri göreli performansını konusunu açıklığa kavuşturmak amacıyla, biz son zamanlarda beş Cas endonucleases – SpCas9, Staphylococcus aureus (SaCas9), Cas9 enzim dan gelen Cas9 enzim sistematik karşılaştırılması gerçekleştirdiyseniz Neisseria meningitidis (NmCas9), Cas12a enzim Acidaminococcus sp. BV3L6 üzerinden (AsCas12a) ve Cas12a enzim (LbCas12a) Lachnospiraceae bakteri ND2006 üzerinden25. Adil bir karşılaştırma için hedef siteleri ve diğer deneysel koşullar aynı kümesini kullanarak çeşitli Cas nükleaz değerlendirildi. Teknoloji kullanıcıları için yararlı bir başvuru olarak hizmet verecek her CRISPR-Cas sistem ayrıca belirlenen çalışma tasarım parametreleri. Burada, daha iyi araştırmacılar CRISPR-CA’larına faydalanmak için etkinleştirme sistemi, biz en iyi genom Mühendisliği (bkz. şekil 1) farklı Cas9 ve Cas12a enzimleri ile için adım adım bir protokol sağlar. Protokol sadece başarılı genom mühendislik sonucu memeli hücrelerinde olasılığını en üst düzeye çıkarmak için de önemli tasarım konuları ama deneysel ayrıntıları içerir.
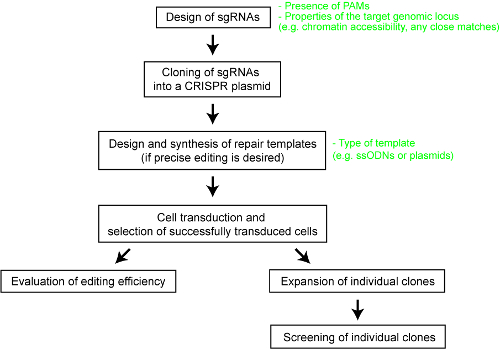
Resim 1 : İnsan hücre hatları düzenlenmiş genom oluşturmak için iş akışının genel bir bakış. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
