


Summary
Presentamos una metodología para establecer los requisitos de polinización de los cultivares de albaricoque (Prunus armeniaca L.) que combinan la determinación de la auto-(in)compatibilidad por microscopía de fluorescencia con la identificación del S-genotipo por análisis PCR.
Abstract
La autoincompatibilidad en Rosaceae está determinada por un sistema de autoincompatibilidad Gametophytic (GSI) que está controlado principalmente por el locus multiallelic S. En el albaricoque, la determinación de las relaciones de auto-e inter-(in)compatibilidad es cada vez más importante, ya que la liberación de un número importante de nuevos cultivares ha dado lugar al aumento de cultivares con requisitos de polinización desconocidos. Aquí, describimos una metodología que combina la determinación de la auto-(in)compatibilidad por polinización manual y microscopía con la identificación del genotipo Spor análisis de PCR. Para la determinación de auto-(in)compatibilidad, las flores en la etapa de globo de cada cultivar se recolectaron en el campo, se polinizaron a mano en el laboratorio, fijadas y teñidas con azul anilina para la observación del comportamiento del tubo de polen bajo la microscopía de fluorescencia. Para el establecimiento de relaciones de incompatibilidad entre cultivares, el ADN de cada cultivar se extrajo de hojas jóvenes y los S-alelos fueron identificados por PCR. S Este enfoque permite establecer grupos de incompatibilidad y dilucidar las relaciones de incompatibilidad entre los cultivares, lo que proporciona una valiosa información para elegir polinizadores adecuados en el diseño de nuevos huertos y para seleccionar padres apropiados en programas de cría.
Introduction
La autoincompatibilidad es una estrategia de floración para prevenir la autopolinización y promover el cruce1. En Rosaceae, este mecanismo está determinado por un sistema de autoincompatibilidad Gametophytic (GSI) que está controlado principalmente por el locus multiallelic S2. En el estilo, el gen RNase codifica el determinante Tylar S-s,un RNase3,mientras que una proteína F-box, que determina el determinante del polen S,está codificada por el gen SFB 4. La interacción de autoincompatibilidad se lleva a cabo a través de la inhibición del crecimiento del tubo de polen a lo largo del estilo evitando la fertilización del óvulo5,6.
En albaricoque, en lasúltimas,dos décadas se ha producido una renovación varietal en todo elmundo. Esta introducción de un número importante de nuevos cultivares, de diferentes programas de cría públicos y privados, ha dado lugar al aumento de cultivares de albaricoque con requisitos de polinización desconocidos8.
Se han utilizado diferentes metodologías para determinar los requisitos de polinización en el albaricoque. En el campo, la autocompatibilidad puede establecerse mediante polinizaciones controladas en árboles enjaulados o en flores emasculadas y,posteriormente,registrar el porcentaje del conjunto de frutas9,10,11,12. Además, se han realizado polinizaciones controladas en el laboratorio mediante el cultivo semi-in vivo de flores y el análisis del comportamiento del tubo de polen bajo microscopía de fluorescencia8,13,14,15,16,17. Recientemente, las técnicas moleculares, como el análisis de PCR y la secuenciación, han permitido caracterizar las relaciones de incompatibilidad basadas en el estudio de los genes RNase y SFB 18,,19. En el caso del albaricoque, se han notificado treinta y tres S-aleles (S1 a S20, S22 a S30, S52, S53, Sv, Sx), incluyendo un alelo relacionado con la autocompatibilidad (Sc)12,18,20,21,22,23,24. Hasta ahora, 26 grupos de incompatibilidad han sido estabilizados en esta especie según el S-genotipo8,9,17,25,26,27. Los cultivares Scon los mismos S-aleles son inter-incompatibles, mientras que los cultivares con al menos un S-allelediferente y, en consecuencia, asignados en diferentes grupos incompatibles, son inter-compatibles.
Para definir los requisitos de polinización de los cultivares de albaricoque, describimos una metodología que combina la determinación de la auto-(in)compatibilidad por microscopía de fluorescencia con la identificación del genotipo Spor análisis de PCR en cultivares de albaricoque. Este enfoque permite establecer grupos de incompatibilidad y dilucidar las relaciones de incompatibilidad entre cultivares.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Determinación de auto-(in)compatibilidad
- Pruebe las flores en el campo. Es necesario recoger las flores en la etapa de globo(Figura 1A), correspondiente a la etapa 58 en la escala BBCH para el albaricoque28,para evitar la polinización previa no deseada.
-
Autonuquinaciones cruzadas y autonúditos en el laboratorio
- Retire las anteras de las flores en la etapa del globo y colóquelas en un pedazo de papel para secar a temperatura de laboratorio.
- Después de 24 h, tamiza los granos de polen usando una malla fina (0,26 mm) (Figura 1B).
- Emascular un grupo de 30 flores en la misma etapa de globo para cada autopolinización y polinización cruzada y colocar los pistilos en espuma florista en agua a temperatura de laboratorio (Figura 1C).
- Polinizar a mano los pistilos con la ayuda de un pincel con polen de flores del mismo cultivar 24 h después de la emasculación. Además, polinizar otro conjunto de pistilos de cada cultivar con polen de flores de un polinizador compatible como control (Figura 1D).
- Después de 72 h, fijar los pistilos en una solución fijativa de etanol/ácido acético (3:1) durante al menos 24 h a 4 oC29. A continuación, deseche el fijador y agregue un 75% de etanol asegurándose de que las muestras estén completamente sumergidas en la solución. Las muestras se pueden conservar en esta solución a 4oC hasta que seutilicen 8,17,30,31,32.
-
Evaluación de la viabilidad del polen a través de la germinación in vitro del polen
- Para preparar el medio de germinación, peso 25 g de sacarosa, 0,075 g de ácido bórico (H3BO3) y 0,075 g de nitrato de calcio (Ca(NO3)2)33.
- Añadir los componentes del medio en 250 ml de agua destilada y disolver por completo.
- Solidificar el medio añadiendo 2 g de agarosa y mezclar remolinos.
- Compruebe el pH del medio utilizando un medidor de pH y ajuste el valor a 7.0 con la solución NaOH o HCl.
- Autoclave la mezcla para esterilizar el medio.
- Después del autoclavado, enfríe el medio y distribúyalo en platos Petri en una capucha de flujo laminar estéril.
- Dispersar los granos de polen de los mismos cultivares utilizados para las polinizaciones controladas en el medio de germinación de polen solidificado y observarlos bajo el microscopio después de 24 h6.
NOTA: Para esterilizar la campana de flujo laminar, limpie la superficie con un 70% de etanol y encienda la lámpara UV durante 10 min. - Almacene los platos de Petri en nevera a 4 oC hasta su uso.
-
Observaciones de microscopía
- Lavar los pistilos tres veces durante 1 h con agua destilada y dejarlos en sulfito sódico al 5% a 4 oC. Después de 24 h, autoclavelos a 1 kg/cm2 durante 10 min en sulfito de sodio para suavizar los tejidos34.
- Coloque los pistilos autoclaves sobre un tobogán de vidrio y, con la ayuda de un bisturí, retire los tricomas alrededor del ovario para obtener una mejor visualización de los tubos de polen. A continuación, aplastar los pistilos con un vaso de cubierta.
- Preparar la mancha azul anilina del 0,1% (v/v): mezclar 0,1 ml de azul anilina en 100 ml de 0,1 N de fosfato potásico tribásico (K3PO4). Aplique una gota de azul anilina sobre las preparaciones para manchar las deposiciones de callosa durante el crecimiento del tubo de polen.
- Observe los tubos de polen a lo largo del estilo mediante un microscopio con epifluorescencia UV utilizando 340-380 filtros de paso de banda y 425 filtros de paso largo.
2. Extracción de ADN
- Muestra 2-3 hojas en el campo en primavera. Se recomienda tomar muestras de las hojas en etapas jóvenes ya que el ADN obtenido es de mayor calidad y niveles más bajos de compuestos fenólicos en comparación con las hojas viejas.
- Extraiga ADN genómico siguiendo los pasos descritos en un kit disponible comercialmente (consulte Tabla de materiales).
- Analizar la cantidad y calidad de las concentraciones de ADN utilizando espectrofotómetro UV-vis (260 nm).
3. IdentificaciónS-allele
-
Configuración de las reacciones DE PCR
- Preparar una dilución de 50 ng/L en agua destilada de cada muestra de extracción de ADN.
- Descongele los reactivos de PCR lentamente y manténgalos sobre hielo. Deje el ADN polimerasa en el congelador hasta que sea necesario.
- Preparar las reacciones de amplificación utilizando las diferentes combinaciones de imprimaciones. Cree la mezcla de reacciones PCR combinando los componentes de la Tabla 1. Vortex la reacción PCR mezcla bien y distribuye el volumen indicado para las diferentes combinaciones de imprimaciones a cada pozo de la placa PCR. A continuación, agregue 1 l de la dilución del ADN en cada pocól.
- Coloque la placa PCR en el termociclador y ejecute el programa PCR correspondiente que se muestra en la Tabla 1.
- Analice los fragmentos amplificados. Hay principalmente dos formas diferentes de analizar los fragmentos amplificados PCR: electroforesis capilar (CE) con imprimaciones con etiqueta fluorescente o como visualización amplicons de electroforesis de gel de agarosa con imprimaciones no etiquetadas.
-
Electroforesis capilar
- Para preparar el tampón de carga, mezcle 35 ml de formamida desionizada con 0,45 ml de tamaño etiquetado estándar. Vórtice el reactivo para mezclar bien, y luego dispensar 35.5 l en el pocódcilo de la placa del lector.
- Agregue 1 l del producto PCR en el pocól. Además, añadir una gota de aceite mineral para evitar la evaporación del agua.
- Prepare la placa de separación añadiendo tampón de separación.
- Utilice el software comercial incluido con el analizador de genes (ver Tabla de materiales). Cree una nueva placa de muestra y guarde los nombres de muestra para todos los pozos de la placa.
- Seleccione el método de análisis. En este caso, desnaturalizar las muestras a 90oC durante 120 s, inyectar a 2,0 kV durante 30 s y separar a 6,0 kV durante 35 min.
- Inserte las dos placas en el analizador genético. Llene la matriz capilar con agua destilada.
- Cargue el gel patentado de poliacrilamida lineal (LPA). Por último, haga clic en Ejecutar.
-
Electroforesis de gel
- Preparar un gel de agarosa al 1% añadiendo 1,5 g de agarosa de grado de biología molecular en 150 ml de tampón de electroforesis de 1% TAE (Tris-acetate-EDTA) (40 mM Tris, ácido acético de 20 mM y EDTA de 1 mM a pH 8.0). Disolver la agarosa por calentamiento por microondas durante 2-3 min.
- Para visualizar el ADN, añada 4 s de una mancha de ácido nucleico (ver Tabla de materiales) y mezcle suavemente.
- Añadir un peine de gel, con suficientes pozos para escaleras, controles y muestras, en una bandeja de gel. Luego, vierta lentamente la mezcla en el centro de la bandeja de gel y evite las burbujas.
- Dejar enfriar el gel durante 30-45 min a temperatura ambiente hasta que el gel se haya solidificado por completo. Introducir el gel en la cámara de electroforesis, quitar el peine de gel y llenar la cámara con suficiente tampón 1x TAE para cubrir el gel.
NOTA: Compruebe la colocación del gel. Los pozos deben colocarse cerca del polo negativo ya que el ADN cargado negativamente migra hacia el cátodo. - Añadir 5 l de tampón de carga (0,1% (v/v) azul bromofenol) a los productos de PCR y mezclar bien.
- Para estimar el tamaño de las bandas, cargue 5 s de la escalera de peso molecular del ADN (ver Tabla de materiales).
- Cargue las muestras en los pozos adicionales del gel.
- Una vez cargadas todas las muestras y la escalera de peso molecular del ADN, ejecute el gel a 90 V durante 1-1,5 h, hasta que la línea de tinte azul esté aproximadamente al 75% de la longitud del gel.
- Visualice las bandas en un transilluminador para los ácidos nucleicos.
-
Electroforesis capilar
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Los estudios de polinización en albaricoque requieren el uso de flores en la etapa tardía del globo un día antes de la antítesis (Figura 1A). Esta etapa se considera la más favorable para la recolección de polen y pistilo, ya que las estructuras florales están casi maduras, pero la dehiscencia de las anteras aún no se ha producido. Esto evita la interferencia del polen no deseado, no sólo de polen de la misma flor sino también de otras flores, ya que los pétalos cerrados impiden la llegada de insectos que llevan polen externo. Los granos de polen se tamizan fácilmente a través de una malla fina(Figura 1B)de las anteras dehisced previamente colocadas en un pedazo de papel durante 24 horas a temperatura ambiente o con un ligero calor extra. Del mismo modo, los pistilos se obtienen de las flores en la etapa del globo después de la eliminación de pétalos, sépalos y estambres con la ayuda de pinzas o uñas (Figura 1C). Los pistilos pueden ser auto- y polinizados cruzados con un cepillo fino (Figura 1D).
Las flores hermafroditas de albaricoque tienen cinco sépalos rojos oscuros, cinco pétalos blancos (Figura 1A), un solo pistilo (Figura 2A) y 25-30 estambres. El pistilo tiene tres estructuras principales: estigma, estilo y ovario. El ovario tiene dos ovulos, y la fertilización de al menos uno de ellos es necesaria para el ajuste de la fruta. Durante la polinización, los insectos, principalmente las abejas, transfieren los granos de polen al estigma (Figura 1A), donde germinan (Figura 2B) dentro de las 24 h después de la polinización. Un tubo de polen se produce a partir de cada grano de polen germinante, que crece a través de las estructuras del pistilo para llegar al ovario después de 3-4 días y fertilizar uno de los dos ovulos después de alrededor de 7 días. En cultivares autoincom incompatibles en los que el alelo S del grano de polen es el mismo que uno de los dos pistilos, el tubo de polen deja de crecer en el estilo superior, evitando la fertilización (Figura 2C). Sin embargo, los tubos de polen de un cultivar compatible, con un alelo S diferente, pueden crecer a través del estilo (Figura 2D), llegar al ovario (Figura 2E) y fertilizar una de las dos óvulos.
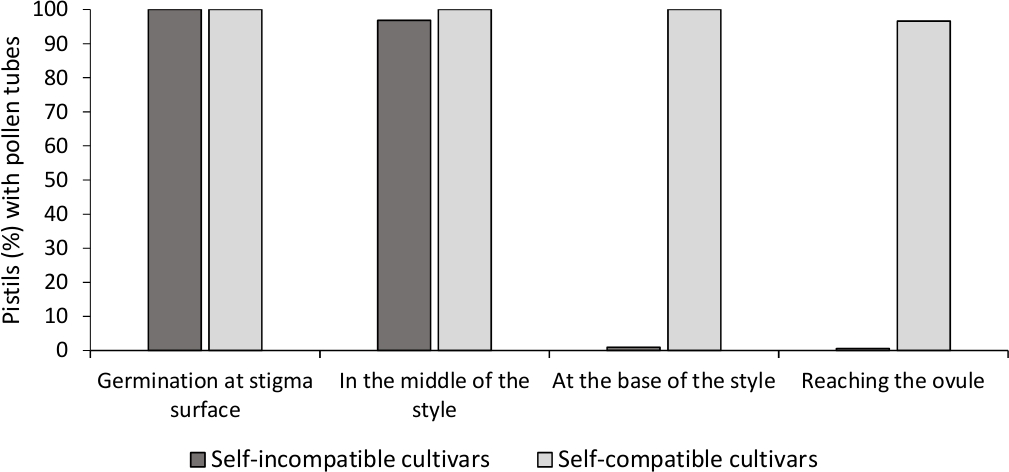
El análisis de la germinación in vitro del polen mostró una buena viabilidad del polen en todos los cultivares analizados aquí, ya que la mayoría de los tubos de polen eran más largos que la longitud del grano de polen después de 24 h en el medio de cultivo. Se observaron granos de polen germinados en la superficie del estigma (Figura 2B) en pistilos de todas las polinizaciones, lo que indica una polinización adecuada (Figura 3).
Para determinar la auto-(in)compatibilidad para cada cultivar, se observó el comportamiento del tubo de polen en auto-y polinizaciones cruzadas realizadas en condiciones controladas por laboratorio bajo microscopía de fluorescencia. El crecimiento del tubo de polen se registró a lo largo del estilo en todos los pistilos examinados. Los cultivares se consideraban autocompatibilidades cuando se detuvo el crecimiento del tubo de polen a lo largo del estilo en la mayoría de los pistilos autopolinizados(Figura 2C, Figura 3)y autocompatibilidad cuando al menos un tubo de polen alcanzó la base del estilo en la mayoría de los pistilos examinados (Figura 2E, Figura 3).
El estudio del S-locus por análisis de PCR permitió caracterizar el genotipo Sde cada cultivar. En primer Slugar, los S-alleles fueron identificados por la amplificación del primer intrón S-RNase utilizando las imprimaciones SRc-F/SRc-R (Tabla 2). El tamaño de los fragmentos amplificados fue analizado por electroforesis capilar (Figura 4A) y se utilizó para clasificar los genotipos analizados en su grupo de incompatibilidad correspondiente (I.G.) (Tabla 3).
Algunos pares de alelos, como S1 y S7 o S6 y S9,mostraron tamaños de fragmento similares para el primer intrón. Así, la diferenciación de estos alelos se realizó amplificando una región del segundo intrón de la RNase con las imprimaciones Pru-C2/PruC4R, SHLM1/SHLM2 y SHLM3/SHLM4 (Tabla 2). La combinación de imprimación PruC2/PruC4R se utilizó para distinguir entre S6 y S9. Para S6,se amplificó un fragmento de 1300 bp, mientras que se observó un fragmento de alrededor de 700 bp para el alelo S9 ( Figura4B,Tabla 3). Las imprimaciones específicas SHLM1/SHLM2 y SHLM3/SHLM4 amplificaron un fragmento de aproximadamente 650 bp en el alelo S1 y 413 bp en el alelo S7 ( Figura4C, Tabla 3).
Las imprimaciones AprFBC8-(F/R) que amplifican las regiones variables V2 y HVb del gen SFB se utilizaron para distinguir alelos Sc y S8, ya que ambos alelos muestran una secuencia RNase idéntica. El alelo S8 mostró un fragmento de PCR de aproximadamente 150 bp mientras que un fragmento de 500 bp correspondía al alelo Sc (Figura 4D,Tabla 3). Una Svez determinado el S-genotipo para todos los cultivares, se asignaron cultivares autoincompatibilidades a sus correspondientes grupos de incompatibilidad en función de sus S-aleles (Tabla 3). S
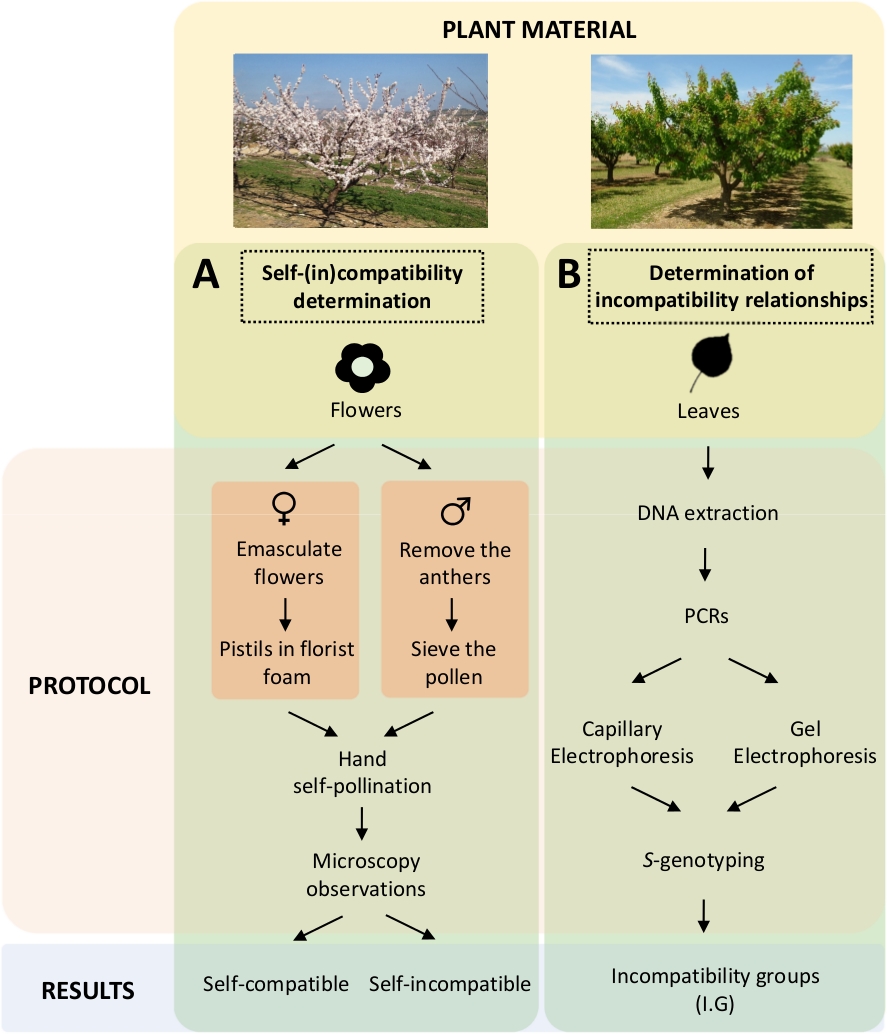
Este enfoque requiere determinar la auto-(in)compatibilidad de cada cultivar mediante autonuquinaciones controladas y entrenaciones en el laboratorio (Figura 5A) concomitantemente con la caracterización del S-genotipopor análisis genético ( Figura5B). Como resultado, se pueden determinar los requisitos de polinización de cada cultivar y las relaciones de incompatibilidad entre los cultivares de albaricoque.

Figura 1. Configuración experimental para la determinación de la auto-(in)compatibilidad en albaricoque.
(A) Flores en la etapa de globo (flechas negras) en el campo. (B) Tamiz de granos de polen utilizando una malla fina. (C) Pistilos colocados sobre espuma florista en agua. (D) Polinización manual de los pistilos con la ayuda de un pincel. Haga clic aquí para ver una versión más grande de esta figura.

Figura 2. Representación diagramamática de las relaciones de incompatibilidad gametofítica en las flores de albaricoque.
(A) En La autoincompatibilidad gametofítica (GSI), los granos de polen compatibles e incompatibles germinan sobre el estigma. El grano de polen lleva uno de los dos S-alelos del genotipo original, en este caso S1 o S2. S Si el S-alleledel grano de polen coincide con uno de los dos S-alelos del pistilo, en este caso S S1S3, el crecimiento del tubo de polen se inhibe en el tercio superior del estilo. (B) Germinación de granos de polen en la superficie del estigma. (C) Tubo de polen detenido en el estilo que indica un comportamiento incompatible. (D) Tubos de polen que crecen a lo largo del estilo. (E) Tubos de polen en la base del estilo que indican un comportamiento compatible. Barras de escala, 100 m. Haga clic aquí para ver una versión más grande de esta figura.

Figura 3. Resultados representativos de la germinación del polen y el crecimiento del tubo de polen a través del estilo para cultivares autocomedidos e incompatibles con sí mismos después de autopolinizaciones.
Porcentaje de pistilos con granos de polen germinando en la superficie del estigma, con tubos de polen a mitad del estilo, en la base del estilo, y llegando al óvulo. Haga clic aquí para ver una versión más grande de esta figura.

Figura 4. Amplificación de fragmentos de PCR utilizando Scinco combinaciones de pares de imprimación para la identificación de S-alleles.
(A) Salida del analizador génica para las imprimaciones SRc-(F/R) que muestran el tamaño de los dos fragmentos amplificados de la primera región de intrón RNase correspondiente a los S-alleles. S (B) Amplificación PCR utilizando las imprimaciones PruC2/PruC4R para la identificación de los alelos S6 y S9. (C) Productos PCR obtenidos utilizando las imprimaciones específicas SHLM1 y SHLM2 para la diferenciación del alelo S1 y SHLM3 y SHLM4 para distinguir el alelo S7. (D) Amplificación PCR con las imprimaciones AprFBC8-(F/R) para identificar alelos Sc y S8. MI: Escalera de ADN de 1 kb. MII: Escalera de ADN de 100 bp. Haga clic aquí para ver una versión más grande de esta figura.

Figura 5. Esquema del diseño experimental para dilucidar las relaciones de compatibilidad entre sí y entre-(in) en cultivares de albaricoque.
(A) Flujo de trabajo de determinación de auto-(in)compatibilidad mediante polinizaciones controladas en el laboratorio. (B) Flujo de trabajo de la identificación del alelo Smediante enfoques moleculares. Haga clic aquí para ver una versión más grande de esta figura.
| PCR Master Mix | Condiciones del termociclor | ||||||
| Componentes | Concentración final | Reacción de 15 l | Paso del ciclo | Temperatura | Hora | Ciclos | |
| 10x NH4 Búfer de reacción | 10x | 1.5 l | Desnaturalización inicial | 94 oC | 3 min | 1 | |
| Solución de 50 mM MgCl2 | 25 mM | 1.2 L | Desnaturalización | 94 oC | 1 min | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 l | Recocido | 55 oC | 1 min | ||
| Primer SRc-F | 10 M | 0,6 l | Extensión | 72 oC | 3 min | ||
| Primer SRc-R | 10 M | 0,6 l | Extensión final | 72 oC | 5 min | 1 | |
| 500 U Taq ADN polimerasa | 0.5 U | 0,2 l | 4 oC | Mantener | |||
| H2O | 8.3 L | ||||||
| Componentes | Concentración final | Reacción de 25 l | Paso del ciclo | Temperatura | Hora | Ciclos | |
| 10x buffer PCR | 10x | 2.5 l | Desnaturalización inicial | 94 oC | 2 min | 1 | |
| 5x Q-solution | 5x | 5 l | Desnaturalización | 94 oC | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 l | Recocido | 55 oC | 2 min | ||
| Primer PruC2 | 10 M | 0,2 l | Extensión | 68 oC | 2 min | ||
| Primer C4R | 10 M | 0,2 l | Desnaturalización | 94 oC | 10 s | 25 | |
| 250 U Taq ADN polimerasa | 10 U | 0,13 l | Recocido | 58 oC | 2 min | ||
| H2O | 15,5 l | Extensión* | 68 oC | 2 min | |||
| Extensión final | 72 oC | 5 min | 1 | ||||
| 4 oC | Mantener | ||||||
| * con 10 s añadido cada ciclo al paso de extensión 68 %C. | |||||||
| Componentes | Concentración final | Reacción de 25 l | Paso del ciclo | Temperatura | Hora | Ciclos | |
| 10x buffer PCR | 10x | 2.5 l | Desnaturalización inicial | 94 oC | 2 min | 1 | |
| 5x Q-solution | 5x | 5 l | Desnaturalización | 94 oC | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 l | Recocido | 62 oC | 1.5 min | ||
| Primer SHLM1 | 10 M | 0,2 l | Extensión | 72 oC | 2 min | ||
| Primer SHLM2 | 10 M | 0,2 l | Extensión final | 72 oC | 5 min | 1 | |
| 250 U Taq ADN polimerasa | 10 U | 0,13 l | 4 oC | Mantener | |||
| H2O | 15,5 l | ||||||
| Componentes | Concentración final | Reacción de 20 l | Paso del ciclo | Temperatura | Hora | Ciclos | |
| 5x búfer PCR | 5x | 4 l | Desnaturalización inicial | 98 oC | 30 s | 1 | |
| dNTP | 2,5 mM | 1.6 l | Desnaturalización | 98 oC | 10 s | 35 | |
| Primer SHLM3 | 10 M | 1 l | Recocido | 51 oC | 30 s | ||
| Primer SHLM4 | 10 M | 1 l | Extensión | 72 oC | 1 min | ||
| 100 U ADN polimerasa | 5 U | 0,2 l | Extensión final | 72 oC | 5 min | 1 | |
| H2O | 12,4 l | 4 oC | Mantener | ||||
| Componentes | Concentración final | Reacción de 25 l | Paso del ciclo | Temperatura | Hora | Ciclos | |
| 10x buffer PCR | 10x | 2.5 l | Desnaturalización inicial | 94 oC | 2 min | 1 | |
| 100 mM dNTP | 2,5 mM | 2 l | Desnaturalización | 94 oC | 30 s | 35 | |
| Primer FBC8-F | 10 M | 1 l | Recocido | 55 oC | 1.5 min | ||
| Primer FBC8-R | 10 M | 1 l | Extensión | 72 oC | 2 min | ||
| 250 U Taq ADN polimerasa | 10 U | 0.125 l | Extensión final | 72 oC | 5 min | 1 | |
| H2O | 17.4 L | 4 oC | Mantener | ||||
Tabla 1. Condiciones de reacción y ciclo para diferentes combinaciones de imprimación utilizadas en este protocolo.
| Imprimaciones | Secuencia | Referencia |
| SRc-F | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTACTCTTAG-3' | 26 |
Cuadro 2. Primers utilizados en este protocolo, secuencia y referencia para la caracterización S-genotipo en Prunus armeniaca.
| Cultivar | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotipo | Grupo de incompatibilidad (I.G) |
| Wonder Cot8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Cuna Mágica8 | 334, 420 | 749 | S2S9 | Xx | |||
| Goldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Ⅳ | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | XVIII | |||
| Apriqueen | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Spring Blush8 | 274, 358 | 150 | S3S8 | XXI |
Cuadro 3. S-genotipado de cultivares de albaricoque con cinco pares de imprimación utilizados en este protocolo y asignación de grupo de incompatibilidad. S En la tabla se muestran los diferentes tamaños de producto de reacción en cadena de lapolimerasa de S-aleles amplificados con imprimaciones SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 y AprFBC8-(F/R).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Tradicionalmente, la mayoría de los cultivares europeos de albaricoque comercial eran autocomprobados36. Sin embargo, el uso de cultivares autoincom incompatibles de América del Norte como padres en programas de cría en las últimas décadas ha dado lugar a la liberación de un número creciente de nuevos cultivares autoincom incompatibles con requisitos de polinización desconocidos7,,8,,37. Por lo tanto, la determinación de las relaciones de auto-e inter-(in)compatibilidad en los cultivares de albaricoque es cada vez más importante. Esto se acentúa en aquellas zonas donde el escalofriante invernal está disminuyendo, ya que las altas variaciones año tras año en la época de floración están impidiendo la coincidencia en la floración de los cultivares y sus polenizadores en muchos casos, especialmente en cultivares con altos requisitos de enfriamiento38. La metodología descrita en el presente documento, combinando la polinización manual, la microscopía y los análisis genéticos ha sido muy útil para determinar la autocompatibilidad de cada cultivar y para establecer sus posibles cultivares polinizadores.
Los requisitos de polinización se pueden determinar mediante experimentos de control de campo en las condiciones de huerto11,,39. Sin embargo, la exposición a factores externos, incluidas las condiciones adversas meteorológicas, puede causar la insuficiencia de polinización10,lo que puede dar lugar a diagnósticos erróneos de autoincompatibilidad. La metodología descrita en este documento permite evaluar la autocompatibilidad con mayor precisión mediante observaciones de microscopía de flores polinizadas a mano en condiciones controladas por laboratorio, evitando la influencia ambiental. Además, este enfoque permite analizar un mayor número de cultivares por año, ya que sólo se requiere un pequeño número de flores en lugar de varios árboles adultos para cada cultivar que se requieren en experimentos de campo40.
Se pueden establecer relaciones de incompatibilidad combinando polinizaciones manuales y microscopía14. Sin embargo, las polinizaciones sólo se pueden realizar durante un corto período durante la temporada de floración en primavera, y se necesitan árboles adultos cerca del laboratorio, ya que la vida útil de las flores recogidas es muy corta. Por lo tanto, el número de relaciones de incompatibilidad que pueden ser analizadas por polinizaciones manuales controladas en cada temporada es muy bajo. La caracterización de los genes codificados por el S-locusha permitido el desarrollo de métodos basados en PCR para el genotipado S-allele18,,41. Este enfoque Sacelera la identificación S-allele ya que no requiere flores, y los experimentos se pueden llevar a cabo con cualquier tejido vegetativo42. Esto amplía el período durante el cual el material vegetal, generalmente hojas jóvenes, se puede recoger43. Además, las hojas pueden ser liofilizadas o congeladas, por lo que el análisis se puede hacer en cualquier época del año, a diferencia de las polinizaciones que sólo se pueden hacer en flores frescas durante la temporada de floración44. Un beneficio adicional es que las hojas se pueden recoger de los árboles jóvenes incluso antes de entrar en la edad de floración, facilitando la recolección de muestras y la obtención temprana de resultados45.
El análisis genético permite una mejor diferenciación de los alelos de autoincompatibilidad ya que proporciona resultados precisos de fragmentos amplificados tamaños21,,46. Hasta la fecha, se han identificado treinta y tres S-aleles en el albaricoque12,18,20,21,22,23,24, que ha permitido establecer 36 grupos de incompatibilidad basados en S-genotipo8,9,17,25,26,27. S Por otro lado, un inconveniente de esta metodología es que diferentes alelos en el mismo tamaño de rango o mutaciones pueden ser identificados erróneamente como el mismo alelo. Por lo tanto, los alelos Sc y S8 son idénticos para la secuencia RNase, pero una inserción de 358-bp se encuentra en el gen SFB de Sc19. Del mismo modo, la primera región de intrón de los alelos S1 y S7 son idénticas y son indistinguibles utilizando las imprimaciones SRc-F/SRc-R. Además, se han encontrado varias homologías, como S6 y S528 o S20 y S55,y S7, S13 (EF062341) y S4617,porque algunos de estos alelos han sido parcialmente secuenciados o por fallos durante la amplificación de PCR y, en consecuencia, se necesita más trabajo para distinguirlos correctamente.
El análisis de PCR y la secuenciación S-RNase son adecuados para establecer relaciones de incompatibilidad mediante la identificación de S-alleles y la asignación de cultivares en su correspondiente SGrupo de Incompatibilidad8,,17,,26,,27. Sin embargo, esta metodología tiene la limitación de impedir la determinación de la auto-(in)compatibilidad para cultivares de albaricoque particulares. La autocompatibilidad (SC) se ha asociado a particularesS-alleles en otras especies de Prunus 47, como la almendra (Sf)48,49 o cereza dulce (S4')50,51. Sin embargo, en el albaricoque, el alelo Sc, que ha sido asociado a SC21,puede identificarse erróneamente como S8, se ha identificado un alelo autocomens incompatible19,,22y posibles mutaciones no vinculadas al locus S, como el M-locus12,52, que confiere SC. Recientemente, Mel M-locus ha sido genotipado usando los marcadores SSR12. Por lo tanto, la identificación genética de SC para genotipos de albaricoque necesita más investigación y, para evitar errores debidos a factores no vinculados al locus S, en este trabajo la caracterización de la auto-(in)compatibilidad se ha determinado también mediante fenotipado el comportamiento de los tubos de polen a través del pistilo de las flores autopolinizadas.
La metodología descrita en el presente documento combina la determinación de la auto-(in)compatibilidad por polinización manual en condiciones de laboratorio con la posterior observación del comportamiento de los tubos de polen en el pistilo de autopolinizaciones controladas bajo la microscopía de fluorescencia y la identificación del genotipo Spor el análisis pcR permite establecer los requisitos de polinización de los cultivares de albaricoque. Esto proporciona una información valiosa para los cultivadores y criadores, ya que permite establecer las relaciones de incompatibilidad entre los cultivares para elegir polinizadores adecuados en el diseño de nuevos huertos, así como para seleccionar padres apropiados para diseñar nuevos cruces en programas de cría de albaricoque.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Los autores no tienen nada que revelar.
Acknowledgments
Esta investigación fue financiada por ministerio de ciencia, Innovación y Universidades-Fondo Europeo de Desarrollo Regional, Unión Europea (AGL2016-77267-R, y AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragón-Fondo Social Europeo, Unión Europea (Grupo Consolidado A12_17R), Fundación Biodiversidad y Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}