Een groot aantal motiliteitsgebeurtenissen in eukaryote cellen worden gemedieerd door de functie van moleculaire motoreiwitten. Deze motoren bewegen langs de cytoskeletale filamenten, actinefilamenten en microtubuli (MT’s) en zetten de chemische energie van ATP-hydrolyse om in kinetische en mechanische krachten die nodig zijn om de biologische beweeglijkheid in cellen te stimuleren. De op MT gebaseerde S. cerevisiae Cin8 is een bipolair, homotetrameer kinesine-5 motoreiwit dat spindel-MT’s uit elkaar schuift1. Cin8 vervult essentiële functies tijdens mitose, in spindelassemblage 2,3,4 en spilverlenging tijdens anafase 5,6,7. Eerder was aangetoond dat Cin8 een bidirectionele motor is, die directionaliteit schakelt onder verschillende experimentele omstandigheden. Onder omstandigheden met een hoge ionische sterkte bewegen enkele Cin8-motoren bijvoorbeeld naar het min-uiteinde van de MT’s, terwijl cin8-motoren in clusters, in meermotorige MT-glijtests en tussen antiparallel MT’s voornamelijk naar de plus-uiteinden van de MT’sbewegen 8,9,10,11,12 . Deze bevindingen waren om verschillende redenen zeer onverwacht. Ten eerste draagt Cin8 zijn katalytische motorische domein op het amino-eindpunt en dergelijke motoren werden eerder verondersteld uitsluitend plus-end gericht te zijn, terwijl Cin8 werd aangetoond dat minus-end gericht was op het niveau van één molecuul. Ten tweede werd aangenomen dat kinesinemotoren unidirectioneel waren, hetzij minus-end of plus-end gericht, terwijl Cin8 bidirectioneel bleek te zijn, afhankelijk van de experimentele omstandigheden. Ten slotte kon, vanwege de MT-oriëntatie op de mitotische spil, de klassieke rol van kinesine-5-motoren in de scheiding van spindelpolen tijdens spindelassemblage en anafase B alleen worden verklaard door hun plus-end gerichte motiliteit op de MT’s die ze1,13 crosslinken. Na de eerste rapporten over de bidirectionaliteit van Cin8, werd aangetoond dat een paar andere kinesinemotoren bidirectioneel 14,15,16 zijn, wat aangeeft dat de bidirectionele beweeglijkheid van kinesinemotoren vaker voorkomt dan eerder werd aangenomen.
Eerder is gemeld dat Cin8 in cellen ook op een bidirectionele manier beweegt8, wat het idee ondersteunt dat de bidirectionele beweeglijkheid van sommige kinesine-5-motoren belangrijk is voor hun intracellulaire functies. Bovendien, aangezien de drie kinesine-5-motoren waarvan werd gemeld dat ze bidirectioneel waren, afkomstig zijn van schimmelcellen, is onlangs een mogelijke rol voor de bidirectionaliteit van kinesine-5-motoren voorgesteld in dergelijke cellen10. Volgens dit model, bij gesloten mitose van schimmelcellen, waarbij de nucleaire enveloppe niet afbreekt tijdens mitose, leveren kinesine-5-motoren de initiële kracht die de spindelpolen uit elkaar scheidt voorafgaand aan de spindelassemblage. Om deze taak uit te voeren, voorafgaand aan de scheiding van de spindelpool, lokaliseren kinesine-5-motoren zich in de buurt van de spilpolen, door hun minus-end gerichte motiliteit op enkele nucleaire MT’s. Eenmaal op deze positie clusteren kinesine-5-motoren, schakelen ze van directionaliteit, vangen en cross-link MT’s van naburige spindelpolen. Vervolgens zorgen kinesine-5-motoren voor de initiële scheiding van de polen door plus-end gerichte motiliteit op de MT’s die ze crosslinken. Volgens dit model zijn zowel minus-end gerichte motiliteit op enkele MT’s als plus-end gerichte motiliteit op cross-linked MT’s tijdens antiparallel glijden vereist voor schimmel kinesine-5 motoren om hun rol te vervullen in spindelassemblage 1,13.
Het algemene doel van de beschreven methode is het verkrijgen van zeer zuivere schimmel GFP-gelabelde kinesine-5 Cin8 en het uitvoeren van motiliteitstests met één molecuul (figuur 1) terwijl de beweeglijkheid van afzonderlijke moleculen en clusters van Cin8 afzonderlijk wordt geanalyseerd. De scheiding tussen afzonderlijke moleculen en clusters is belangrijk omdat een van de factoren waarvan is aangetoond dat ze de directionaliteit van Cin8 beïnvloeden, de accumulatie in clusters op de MT’s 10,12 is. Alternatieve beweeglijkheidstests, zoals de MT-oppervlakteglijdende en MT-glijdende assays, geven geen informatie over de activiteit van enkelvoudige motoreiwitten17,18. De robuuste motiliteits- en analysemethoden met één molecuul die hier worden beschreven, zijn met succes toegepast om verschillende aspecten van kinesine-5-motoren, Cin8 en Kip1 10,11,12,14,19,20 te karakteriseren.
Hier wordt een gedetailleerd protocol gepresenteerd voor Cin8-overexpressie en -zuivering, polymerisatie van MT’s en de motiliteitstest met één molecuul. Verder worden ook de analyses beschreven om onderscheid te maken tussen afzonderlijke moleculen en clusters van Cin8, en om enkelvoudige motor- en clustersnelheden te bepalen door middel van gemiddelde verplaatsing (MD) en gemiddelde kwadratische verplaatsing (MSD). Dit protocol is bedoeld om onderzoekers te helpen alle stappen van de procedures te visualiseren en te helpen bij het oplossen van problemen met dit soort testen.
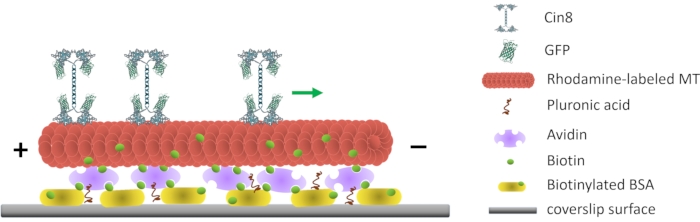
Figuur 1: Schematische weergave van de motiliteitstest met één molecuul. Gebiotinyleerde fluorescerende MT’s zijn bevestigd aan het glazen oppervlak, gecoat met Avidin dat interageert met het aan het oppervlak gehechte biotinylated-BSA. De groene pijl vertegenwoordigt de bewegingsrichting van enkele Cin8-moleculen onder omstandigheden met een hoge ionische sterkte. +/- de polariteit van het MT vertegenwoordigen. Klik hier om een grotere versie van deze figuur te bekijken.
