真核細胞内の多数の運動性事象は、分子運動タンパク質の機能によって媒介される。これらのモーターは、細胞骨格フィラメント、アクチンフィラメント、および微小管(MT)に沿って移動し、ATP加水分解の化学エネルギーを細胞内の生物学的運動性を駆動するために必要な運動学的および機械的力に変換する。MTベースのS. cerevisiae Cin8は、双極性のホモ四量体キネシン-5モータータンパク質であり、スピンドルMTを架橋してスライドさせる1。Cin8は、有糸分裂中、紡錘体アセンブリ2,3,4および分裂後期5,6,7中の紡錘体伸長において、本質的な機能を果たす。これまで、Cin8は双方向モータであり、異なる実験条件下で方向性を切り替えることが実証されていました。例えば、高いイオン強度条件下では、単一のCin8モータはMTのマイナス端に向かって移動するが、クラスタでは、マルチモータMTグライダーアッセイでは、および反並列MT間では、Cin8モータは主にMTのプラス端に向かって移動する8,9,10,11,12 .これらの発見は、いくつかの理由により非常に予想外であった。第一に、Cin8はアミノ末端に触媒モータードメインを持ち、そのようなモーターは以前はもっぱらプラスエンド指向性であると考えられていたが、Cin8は単一分子レベルでのマイナス末端指向性であることが示された。第二に、キネシンモーターはマイナス端またはプラス端のいずれかの単方向であると考えられていたが、Cin8は実験条件に応じて双方向であることが示された。最後に、有糸分裂スピンドルにおけるMT配向のために、スピンドルアセンブリ中のスピンドル極と分裂後期Bの分離におけるキネシン-5モータの古典的な役割は、それらが架橋するMT上のプラスエンド指向性によってのみ説明できた1,13。Cin8の双方向性に関する最初の報告に続いて、他のいくつかのキネシンモーターが双方向であることが実証された14,15,16、キネシンモーターの双方向運動性は、以前に信じられていたよりも一般的である可能性があることを示している。
細胞内では、Cin8も双方向で移動することが以前に報告されており8、いくつかのキネシン5モーターの双方向運動性が細胞内機能にとって重要であるという考えを支持している。また、双方向であることが報告された3つのキネシン-5モーターは真菌細胞由来であることから、このような細胞10においてキネシン-5モーターの双方向性に対する役割の可能性が最近提案されている。このモデルによると、有糸分裂中に核エンベロープが破壊されない真菌細胞の閉鎖有糸分裂では、キネシン-5モーターは、紡錘体組み立ての前に紡錘極を分離する初期力を提供する。このタスクを実行するために、主軸極分離の前に、キネシン-5モータは、単一の核MT上のマイナス端指向運動性によって、主軸極の近くに局在する。この位置に着くと、キネシン-5モーターはクラスタリングし、方向を切り替え、キャプチャし、隣接するスピンドルポールからMTをクロスリンクします。続いて、キネシン-5モータは、それらが架橋するMT上のプラスエンド指向性運動性によって極の初期分離を提供する。このモデルでは、真菌キネシン-5モータがスピンドルアセンブリでその役割を果たすために、単一MT上のマイナス端指向性運動性と、逆平行摺動中の架橋MT上のプラスエンド指向性運動性の両方が必要である1,13。
記載された方法の全体的な目標は、高純度の真菌GFPタグ付きキネシン-5 Cin8を取得し、単一分子およびCin8のクラスターの運動性を別々に分析しながら、単一分子運動性アッセイ(図1)を行うことである。単一分子とクラスターの間の分離は、Cin8の方向性に影響を与えることが実証された要因の1つがMTs10,12上のクラスターへの蓄積であるため、重要である。MT表面滑空およびMT摺動アッセイなどの代替運動性アッセイは、単一モータータンパク質の活性に関する情報を提供しない17、18。ここで説明する堅牢な単一分子運動性アッセイおよび分析方法は、キネシン-5モーター、Cin8およびKip1 10、11、12、14、19、20のさまざまな側面を特徴付けるために首尾よく適用されています。
ここでは、Cin8の過剰発現および精製、MTの重合、および単一分子運動性アッセイについて、詳細なプロトコールを提示する。さらに、Cin8の単一分子とクラスターを区別し、平均変位(MD)および平均二乗変位(MSD)分析によって単一モーター速度およびクラスター速度を決定するための分析も記載されている。このプロトコルは、研究者が手順のすべてのステップを視覚化し、このタイプのアッセイのトラブルシューティングを支援することを目的としています。
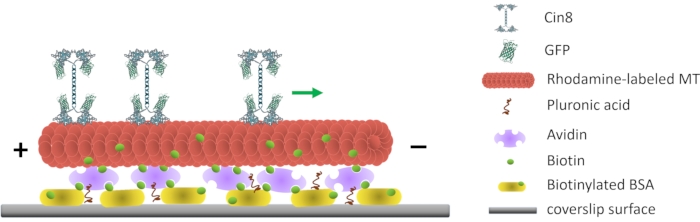
図1:一分子運動性アッセイの概略図。 ビオチン化蛍光MTはガラス表面に付着し、表面結合ビオチン化BSAと相互作用するアビジンでコーティングされている。緑色の矢印は、高イオン強度条件下での単一のCin8分子の移動方向を表す。+/- MTの極性を表します 。この図の拡大版を表示するには、ここをクリックしてください。
