The protocol in the present study has been developed to determine the optimal experimental conditions for measuring physiological phenomena observed in the vascular responses of isolated IPA preparations. The pilot experiments were performed to describe the potential outcomes that aid the understanding of the vascular effects and mechanistic basis of the vasorelaxant action of the plant extract, as follows.
Vasorelaxant effect of the plant extract
As shown in Figure 6A,B, in endothelium-intact IPA (E+), the plant extract elicited a concentration-dependent response of vasorelaxation (EC50 = 66.88 µg/mL, Figure 6C). Eradication of endothelium (E-) profoundly reduced the vasorelaxation induced by the plant extract (p < 0.01), as reflected by the increase in the EC50 by 2.2-fold (E-, EC50 = 150.60 µg/mL, Figure 6C). The vehicle, DMSO, did not possess any effect. Thus, the plant extract produced vasorelaxation mainly via an endothelium-dependent pathway and partially via an endothelium-independent pathway.
Mechanism of vasorelaxant action of the plant extract via endothelium-dependent pathways
As shown in Figure 7, utilizing L-NAME for the inhibition of eNOS (Figure 7B) and the combination of apamin plus charybdotoxin for blocking EDHF (Figure 7D) evidently decreased the vasorelaxant response to the plant extract. This shifted the concentration-response curve to the right and increased the EC50 without altering the Emax values. On the contrary, indomethacin (a COX inhibitor) (Figure 7C), showed no effect on the vasorelaxant response to the plant extract.
Characterizing the role of K+ channels in vasorelaxant action of the plant extract
In the endothelium-denuded IPA rings, KCa channel blocker (iberiotoxin) decreased the vasorelaxant response to the plant extract (Figure 8C), while blockers of Kv (4-AP) or KATP (glibenclamide) channels did not modify the vasorelaxation induced by the plant extract (Figure 8B,D).
Mechanism of vasorelaxant action of the plant extract via extracellular Ca2+ influx inhibition
To investigate whether the mechanism of vasorelaxant action of the plant extract involved extracellular Ca2+ influx inhibition, the vasoconstriction of endothelium-denuded IPA rings was evoked by 1 x 10−5-1 x 10−2 M CaCl2 in Ca2+-free Krebs solution incorporated with 80 mM K+ to activate the VOCCs (Figure 9A,B). Pre-incubation with the plant extract (68 µg/mL, EC50 value) inhibited CaCl2-induced contraction (p < 0.001 vs. vehicle).
Mechanism of vasorelaxant action of the plant extract via inhibition of intracellular Ca2+ release from the SR
To examine whether the release of intracellular Ca2+ from the SR played any role in the vasorelaxant effect, the endothelium-denuded IPA rings were pre-incubated with Ca2+-free Krebs solution, followed by the addition of PE (1 x 10−5 M), rendering a transient contraction (Figure 10A). Then, in the same IPA ring, this experiment was replicated in the presence of either vehicle or plant extract. In comparison to the vehicle, the plant extract significantly reduced (p < 0.001) the contraction induced by PE (Figure 10B).
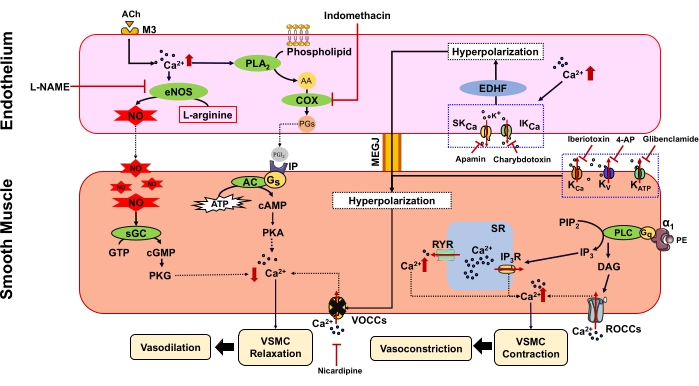
Figure 1: Vascular tone regulation via endothelium-dependent and -independent pathways. AA = Arachidonic acid, ACh = Acetylcholine, AC = Adenylyl cyclase, ATP = Adenosine 5'-triphosphate, cAMP = Cyclic adenosine monophosphate, cGMP = Cyclic guanosine monophosphate, COX = Cyclooxygenase, DAG = Diacylglycerol, EDHF = Endothelium-derived hyperpolarizing factor, eNOS = Endothelial nitric oxide synthase, Gq = G-protein type q, Gs = G-protein type s, GTP = Guanosine triphosphate, IP = Prostacyclin receptor, IP3 = Inositol 1, 4, 5 trisphosphate, IP3R = IP3 receptor, IKCa = Intermediate conductance Ca2+-activated K+ channel, KV = Voltage-gated potassium channels, KATP = ATP-sensitive potassium channels, KCa = Large conductance Ca2+-activated K+ channels, M3 = Muscarinic receptor, MEGJ = Myoendothelial gab junction, NO = Nitric oxide, PE = Phenylephrine, PGI2 = Prostacyclin, PGs = Prostaglandins, PIP2 = Phosphatidylinositol 4,5 bisphosphate, PKA = Protein kinase A, PKG = Protein kinase G, PLA2 = Phospholipase A2, PLC = Phospholipase C, ROCCs = Receptor-operated Ca2+ channels, RYR = Ryanodine receptor, sGC = Soluble guanylyl cyclase, SKCa = Small conductance Ca2+-activated K+ channel, SR = Sarcoplasmic reticulum, VOCCs = Voltage-operated Ca2+ channels. Please click here to view a larger version of this figure.
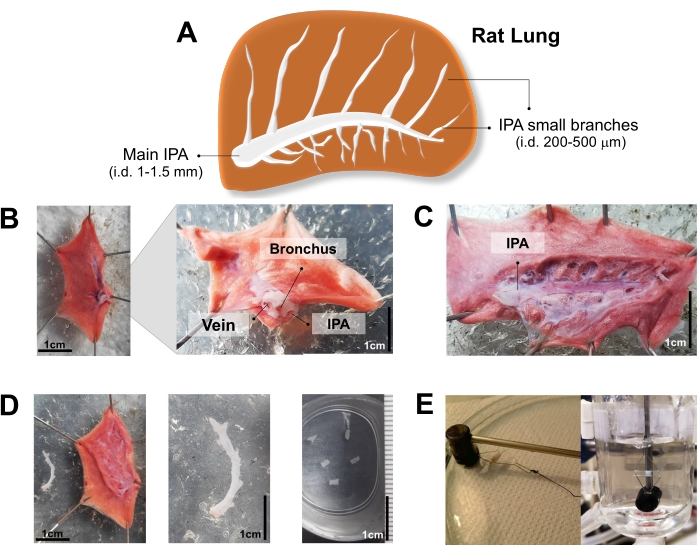
Figure 2: Key steps of the rat intrapulmonary artery (IPA) isolation. (A) The picture depicts rat lung with IPA. (B) Dissection of the medial side/root of the lung facing up. (C) Visualized main IPA after removing the veins and bronchia. (D) Isolated IPA. (E) The IPA rings were mounted onto a pair of stainless-steel wires for a vascular response study using the organ bath technique. Please click here to view a larger version of this figure.
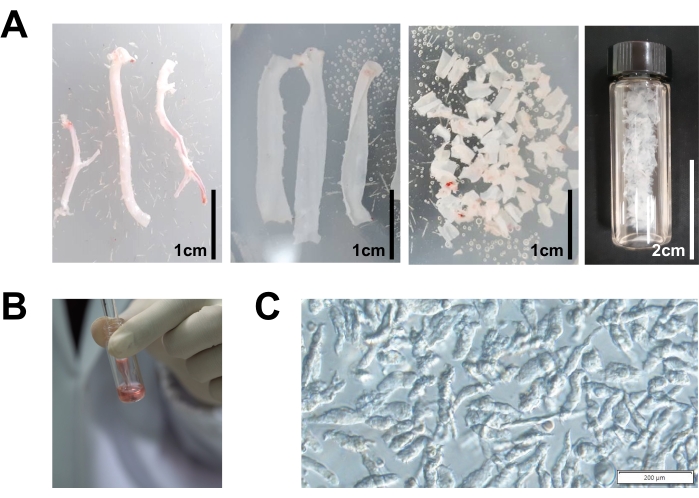
Figure 3: Key steps of the IPA vascular smooth muscle cells (VSMCs) isolation. (A) Isolated IPA was cut into small strips and immersed in dissociation medium (DM). (B) Trituration of vascular strips to isolate VSMCs. (C) Isolated VSMCs after gentle trituration. Please click here to view a larger version of this figure.
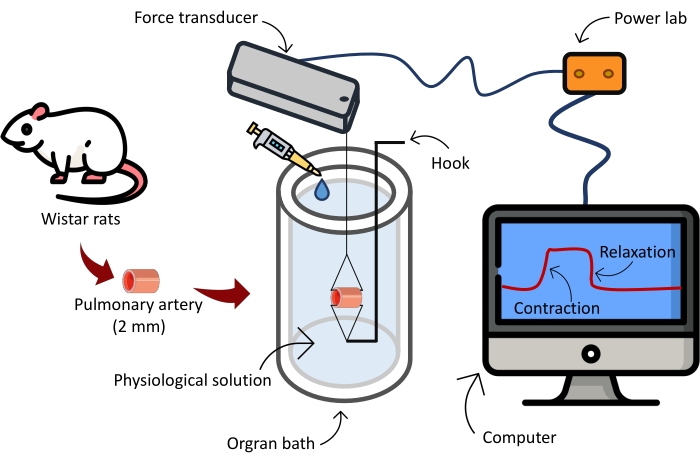
Figure 4: Schematic illustration of the equipment used to test the vascular reactivity. Please click here to view a larger version of this figure.
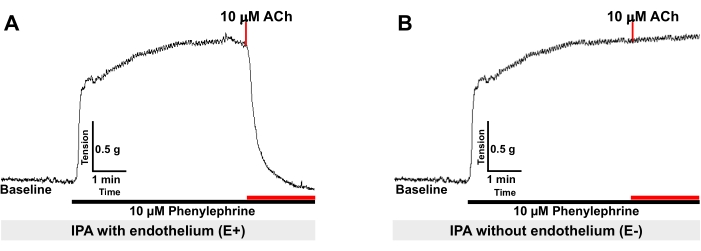
Figure 5: Representative record showing vasorelaxation of IPA rings precontracted with 10 µM PE by 10 µM acetylcholine (ACh). (A) Endothelium-intact ring (E+) and (B) endothelium-denuded (E-) ring. Please click here to view a larger version of this figure.
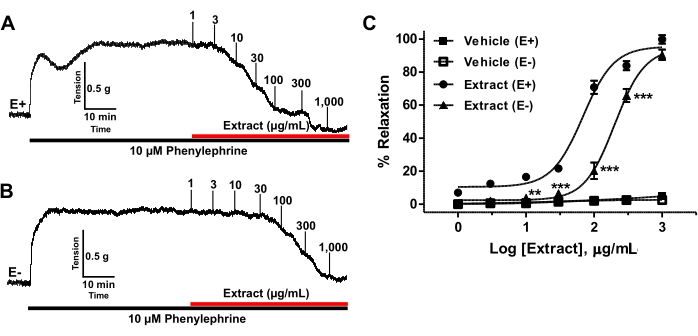
Figure 6: Vasorelaxation of IPA by the plant extract. (A) Representative record showing vasorelaxation of IPA rings by the plant extract (1-1,000 µg/mL) pre-contracted with 10 µM PE in endothelium-intact rings (E+) and (B) endothelium-denuded (E-) rings. (C) Concentration-response curves of vasorelaxation induced by the plant extract in IPA rings (E+, n = 6 and E-, n = 6). Vasorelaxation is expressed as a percentage of the contraction induced by PE. All data are expressed as mean ± SEM. **p < 0.01, ***p < 0.001 compared with vehicle. Please click here to view a larger version of this figure.
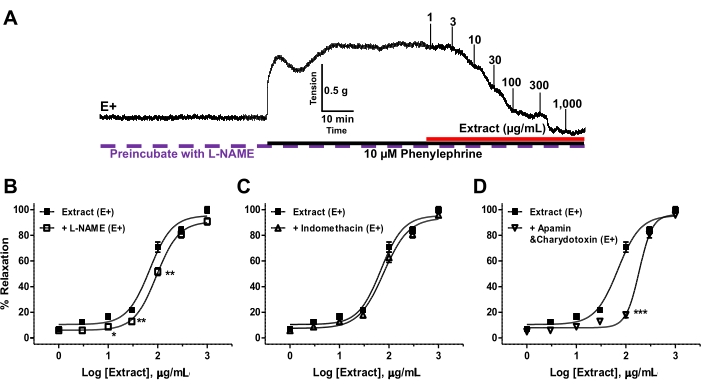
Figure 7. The mechanisms of the plant extract-induced IPA vasorelaxation via the endothelium-dependent pathway. (A) Representative record showing vasorelaxation by the plant extract (1-1,000 µg/mL) of endothelium-intact IPA rings (E+) preincubated with L-NAME (eNOS inhibitor) and precontracted with 10 µM PE. (B–D) Concentration-response curves of the plant extract-induced vasorelaxations of endothelium-intact (E+) IPA rings precontracted with PE and preincubated with inhibitors of various endothelial signaling pathways, including (B) 100 µM L-NAME, (C) 10 µM indomethacin, or (D) 0.1 µM apamin plus 0.1 µM charybdotoxin. Values are means ± SEM. (n = 6). **p < 0.01, ***p < 0.001. Please click here to view a larger version of this figure.
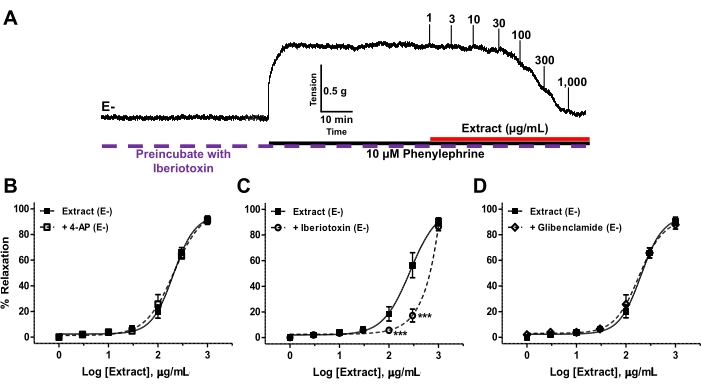
Figure 8. Effect of K+ channel blockers on the plant extract-induced IPA vasorelaxation. (A) Representative record showing vasorelaxation by the plant extract (1-1,000 µg/mL) of endothelium-denuded (E-) IPA rings incubated with 4-AP (KV channel blocker) and precontracted with 10 µM PE. (B–D) Concentration-response curves of the plant extract-induced vasorelaxation of endothelium-denuded (E-) IPA rings precontracted with PE and preincubated with various K+ channel blockers, including (B) 1 mM 4-AP, (C) 10 µM glibenclamide, or (D) 30 nM iberiotoxin. Values are means ± SEM. (n = 6). ***p < 0.001. Please click here to view a larger version of this figure.
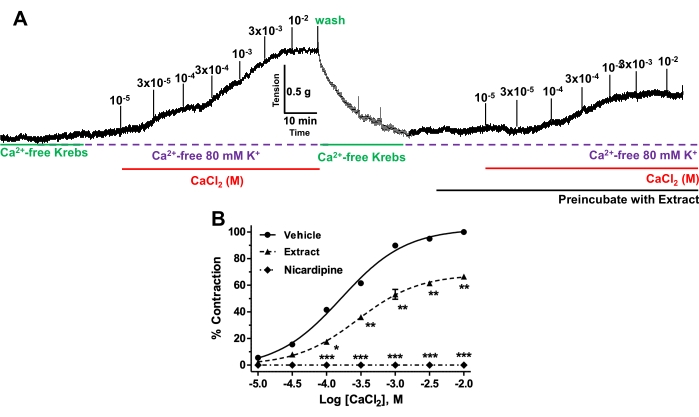
Figure 9. Effect of the plant extract on extracellular Ca2+ influx. (A) Representative records showing CaCl2-induced contraction of IPA rings in the absence (control) or presence of the plant extract. IPA rings were bathed in Ca2+free high K+-solution (80 mM) containing 10 mM EGTA and the contraction evoked by a cumulative concentration of CaCl2 was measured. This protocol was then repeated alone (control, n = 6) or in the presence of the plant extract (n = 6). (B) Concentration-response curves for CaCl2-induced contraction of IPA rings in the absence (control) or presence of the plant extract or 1 µM nicardipine (L-type Ca2+ channel blocker). The CaCl2-induced contraction was calculated as percentage of the maximum contraction recorded from the first CaCl2 application and expressed as mean ± SEM. *p < 0.05, ***p < 0.001 compared with nicardipine. Please click here to view a larger version of this figure.
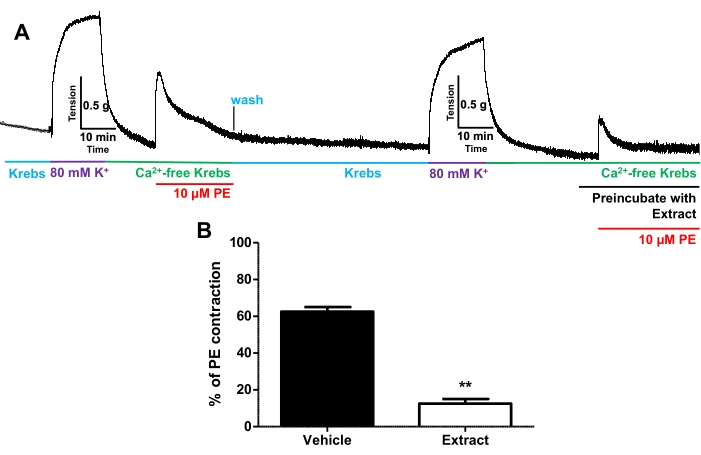
Figure 10. Effect of the plant extract on Ca2+ release from the sarcoplasmic reticulum (SR). (A) Representative record showing phenylephrine (PE)-induced contraction of IPA rings by Ca2+ release from the SR of endothelium-denuded IPA rings in the presence of DMSO (control) and 10 µM of plant extract. The data is percentage contraction to 10 µM PE-induced contractions compared to contractions produced by the initial protocol without the plant extract. Values are means ± SEM. **p < 0.01 compared with vehicle. Please click here to view a larger version of this figure.
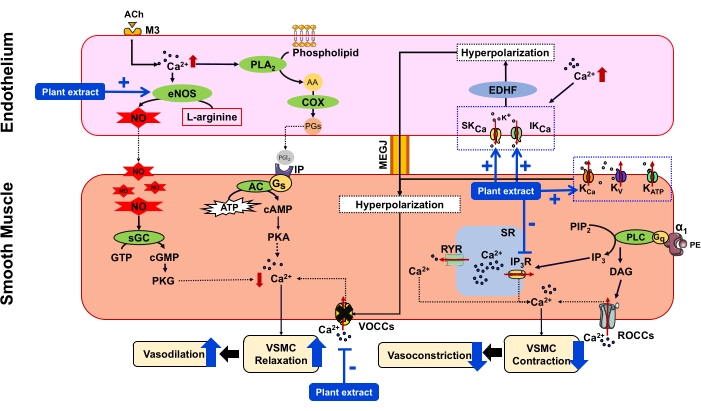
Figure 11: Proposed mechanism of vasodilator action of the plant extract on rat intrapulmonary artery via endothelium-dependent and -independent pathways AA = Arachidonic acid, ACh = Acetylcholine, AC = Adenylyl cyclase, ATP = Adenosine 5'-triphosphate, cAMP = Cyclic adenosine monophosphate, cGMP = Cyclic guanosine monophosphate, COX = Cyclooxygenase, DAG = Diacylglycerol, EDHF = Endothelium-derived hyperpolarizing factor, eNOS = Endothelial nitric oxide synthase, Gq = G-protein type q, Gs = G-protein type s, GTP = Guanosine triphosphate, IP = Prostacyclin receptor, IP3 = Inositol 1, 4, 5 trisphosphate, IP3R = IP3 receptor, IKCa = Intermediate conductance Ca2+-activated K+ channel, KV = Voltage-gated potassium channels, KATP = ATP-sensitive potassium channels, KCa = Large conductance Ca2+-activated K+ channels, M3 = Muscarinic receptor, MEGJ = Myoendothelial gab junction, NO = Nitric oxide, PE = Phenylephrine, PGI2 = Prostacyclin, PGs = Prostaglandins, PIP2 = Phosphatidylinositol 4,5 bisphosphate, PKA = Protein kinase A, PKG = Protein kinase G, PLA2 = Phospholipase A2, PLC = Phospholipase C, ROCCs = Receptor-operated Ca2+ channels, RYR = Ryanodine receptor, sGC = Soluble guanylyl cyclase, SKCa = Small conductance Ca2+-activated K+ channel, SR = Sarcoplasmic reticulum, VOCCs = Voltage-operated Ca2+ channels. Please click here to view a larger version of this figure.
