التربة هي بيئة متنوعة بشكل استثنائي تحتوي على وفرة من الكائنات الحية الدقيقة التي لها دور أساسي في دورات الكربون والفوسفور 1,2. الفطريات الخيطية هي عنصر رئيسي في العديد من النظم الإيكولوجية كمتحللة للمواد العضوية وغير العضوية ويمكن أن تعزز تغذية النباتات من خلال تشكيل علاقات تكافلية 3,4. داخل التربة ، تتفاعل الفطريات ديناميكيا مع العديد من الميكروبات مثل الفطريات الأخرى5 والبكتيريا6 والفيروسات7 والديدان الخيطية8. هذه التفاعلات لها عواقب وخيمة على صحة التربة والنبات. ومع ذلك، وبسبب الافتقار إلى النظم التجريبية المناسبة القادرة على تصوير الكائنات الحية الدقيقة المتفاعلة بدقة عالية، لا يزال العديد منها غير محدد.
وللتحقيقات المتعلقة بالتفاعلات البكتيرية الفطرية والتفاعلات الفطرية والفطرية تطبيقات قيمة في طائفة من المجالات، بما في ذلك مضادات الميكروبات في الطب وعوامل المكافحة البيولوجية في الزراعة. على سبيل المثال ، ينتج الفطر Coprinopsis cinerea الببتيد copsin، الذي ثبت أنه يظهر نشاطا مضادا للبكتيريا ضد العامل الممرض البشري Listeria monocytogenes9. وبالمثل ، يستخدم المركب المشتق من الفطريات ، griseofulvin ، على نطاق واسع كعلاج للعدوى الفطرية البشرية ، بالإضافة إلى أنه قادر على تثبيط نمو الفطريات المسببة للأمراض النباتية Alternaria solani10,11. كما ثبت أن العديد من سلالات بكتيريا Bacillus subtilis التي تعيش في التربة هي عوامل فعالة للمكافحة الحيوية للنبات الفطري الممرض Rhizoctonia solani12,13. ومع ذلك، ونظرا للقيود المرتبطة بالمنهجيات التقليدية، فإن مؤسسات التمويل الدولية ومؤسسات التمويل الأجنبي غير مفهومة بشكل جيد على مستوى الخلايا المفردة.
عادة ما تستكشف الدراسات التقليدية BFIs و FFIs على النطاق الكلي باستخدام ألواح أجار مع نوعين أو أكثر في المواجهة. يتم تقييم تفاعلها عن طريق قياس معدلات النمو وإنتاج الأيض للأنواع المواجهة14،15،16 ؛ ومع ذلك ، يتم حل هذه المنهجية فقط على مستوى المستعمرة. لدراسة التفاعلات على المستوى الخلوي ، يمكن زراعة اللقاحات البكتيرية والفطرية على شرائح المجهر الزجاجية المغلفة بالأجار والتي يتم تصويرها بعد ذلك تحت المجهر17. ومع ذلك ، قد يكون من الصعب اتباع hypha واحد باستخدام شرائح المجهر بسبب عدم وجود حبس ، مما يعني أنه من الصعب الحصول على صور الفاصل الزمني. علاوة على ذلك ، فإن فرصة حصر الكائنات الحية الدقيقة الأخرى مكانيا داخل مناطق محددة من الميسيليوم الفطري أو إنشاء بيئات كيميائية محددة يمكن إزعاجها ، على سبيل المثال ، غير ممكنة في مثل هذه الإعدادات. تضيف طبيعة “الصندوق الأسود” للتربة أيضا إلى تعقيد دراسة التفاعلات الفطرية الميكروبية على مستوى الخلايا المفردة18. من خلال مراقبة الأنواع المتفاعلة بعيدا عن التنوع المذهل لميكروبيوم التربة ، يمكن تقييم الطريقة الدقيقة التي يتفاعل بها الأعضاء الأفراد. وبالتالي ، هناك حاجة مستمرة إلى منصات متعددة الاستخدامات تمكن من التصوير عالي الدقة وخلية واحدة ل BFIs و FFIs.
توفر تقنيات الموائع الدقيقة ، ما يسمى بأنظمة المختبر على رقاقة ، منصة مثالية لدراسة BFIs و FFIs على مستوى الخلايا المفردة. تم اعتماد مجال الموائع الدقيقة ، الناشئة عن التقنيات المطورة للتحليل الكيميائي والإلكترونيات الدقيقة ، من قبل العلوم البيولوجية19. تنظم تقنيات الموائع الدقيقة كميات صغيرة من السوائل داخل شبكة مخصصة من القنوات المصغرة ، لها بعد واحد على الأقل على مقياس الميكرومتر ، ويتوسع استخدامها في الأبحاث البيولوجية20. على وجه الخصوص ، تم تطوير أجهزة الموائع الدقيقة لدراسة نمو الفطريات الخيطية21،22،23،24،25،26،27،28،29،30. إحدى فوائد استخدام هذه التكنولوجيا هي أن حبس hyphae وتوزيع العناصر الغذائية داخل القنوات الدقيقة يشبه إلى حد كبير بنية بيئة التربة أكثر من طرق الأجار التقليدية31. في الآونة الأخيرة ، تم استخدام منصات الموائع الدقيقة للتحقيق في التفاعلات بين العدلات البشرية ومسببات الأمراض الفطرية32 ، والبكتيريا وجذور النباتات 33 ، وكذلك الفطريات والديدان الخيطية34,35.
واحدة من المزايا العديدة لاستخدام الموائع الدقيقة لدراسة التفاعلات الميكروبية تشمل التحكم المحدد في بيئة القنوات الدقيقة. على سبيل المثال ، يمكن استغلال أنظمة التدفق الرقائقي لتوليد تدرجات تركيز محددة ، وهو أمر مفيد بشكل خاص عند فحص التاكسي الكيميائي البكتيري36. ميزة أخرى هي أن الطبيعة الشفافة ل poly (dimethylsiloxane) (PDMS) ، وهو بوليمر مطاطي غير مكلف ومتوافق حيويا يستخدم عادة في تصنيع أجهزة الموائع الدقيقة ، يسهل التصوير عالي الدقة للخلايا المفردة باستخدام المجهر الساطع والفلوري37. وبالمثل ، فإن حبس الميكروبات داخل القنوات الدقيقة يعني أنه يمكن إجراء تجارب الفاصل الزمني التي تتعقب الخلايا المفردة ، مما يسمح بتسجيل الاستجابات الخلوية الفردية وتحديدها كميا37. وأخيرا، بما أنه يمكن تصميم أجهزة الموائع الدقيقة لتكون سهلة الاستخدام، يمكن استخدامها بسهولة من قبل غير الخبراء38.
ومن المهم تعزيز المعرفة بالتفاعلات بين الكائنات الحية الدقيقة التي تعيش في التربة لتحسين ممارسات الإدارة المستدامة للنظم الإيكولوجية التي تحافظ على التنوع البيولوجي وللتخفيف من أثر تغير المناخ على البيئات الأرضية39. وبالتالي ، فإن تطوير أدوات جديدة للموائع الدقيقة أمر أساسي لتوسيع فهم الفطريات وتفاعلاتها على المستوى الخلوي. وسيركز البروتوكول هنا على جهازين من أجهزة الموائع الدقيقة المنتجة لدراسة BFIs40 و FFIs41 كما هو موضح في الشكل 1.
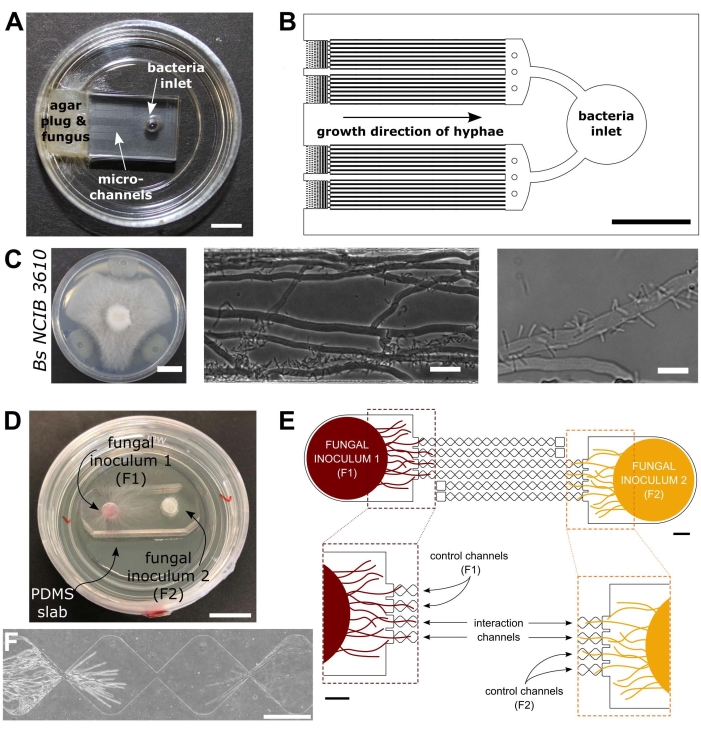
الشكل 1: التمثيل البصري والتخطيطي لأجهزة التفاعل البكتيري الفطري (BFI) والتفاعل الفطري الفطري (FFI). (أ) صورة لجهاز BFI. يتم وضع قابس mycelial عند مدخل أحد طرفي القنوات الدقيقة للسماح بنمو hyphal في الجهاز. المدخل البكتيري في الطرف المعاكس. شريط المقياس = 5 مم (B) نظرة عامة تخطيطية على جهاز BFI ، تصور موضع المداخل البكتيرية واتجاه نمو الواصلة من خلال القنوات الدقيقة التفاعلية. يبلغ عمق القنوات 10 ميكرومتر وعرضها 100 ميكرومتر وطولها 7 مم ، مع 28 قناة مراقبة في المجموع. (ج) فحص المواجهة على صفيحة أجار بين Coprinopsis cinerea و Bacillus subtilis NCIB 3610 ، شريط المقياس = 20 مم (يسار). صور مجهرية تظهر التفاعل بين C. cinerea و B. subtilis NCIB 3610 داخل القناة الدقيقة (الوسطى واليمنى) ، أي التعلق القطبي للبكتيريا ب hyphae الفطرية. شريط المقياس = 25 ميكرومتر (وسط) و 10 ميكرومتر (يمين). (د) صورة لجهاز FFI مرتبط بطبق بتري ذو قاع زجاجي ، ملقح مزدوج بسدادات mycelial. شريط المقياس = 1 سم (E) نظرة عامة تخطيطية لجهاز FFI. يتم إدخال اثنين من سدادات التطعيم الفطرية في المداخل في أي من طرفي الجهاز ، مما يسمح بالاستكشاف الواصلي للقنوات الدقيقة. ترتبط قنوات التحكم بمدخل فطري واحد فقط ولها قناة مسدودة ، مما يمنع التفاعلات بين فطريات الاختبار. تربط قنوات التفاعل كلا من المداخل الفطرية وتسمح بالتفاعلات الواصلة بين مواضيع الاختبار داخل القناة الدقيقة. تتكون كل قناة تفاعل من 18 قسما على شكل ماس ، يبلغ طولها الإجمالي 8.8 مم (490 × 430 ميكرومتر لكل ماسة) ، وعمقها 10 ميكرومتر ، ولها منطقة ربط بين كل ماسة تبلغ 20 ميكرومتر. يتم تكرار أنواع القنوات ، أشرطة المقياس = 1 مم. (F) منطقة التفاعل بين جبهتين واصليتين تقتربان ، تنمو من طرفي نقيض من قناة التفاعل المترابطة. صورة مجهرية لتباين الطور ، شريط مقياس = 250 ميكرومتر. تم تعديل اللوحات الواردة في هذا الشكل من Stanley et al., 2014 (A-C)40 and Gimeno et al., 2021 (D-F)41. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
