Le sol est un environnement exceptionnellement diversifié contenant une abondance de micro-organismes qui jouent un rôle déterminant dans les cyclesdu carbone et du phosphore 1,2. Les champignons filamenteux sont une composante majeure de nombreux écosystèmes en tant que décomposeurs de matière organique et inorganique et peuvent améliorer la nutrition des plantes grâce à la formation de relations symbiotiques 3,4. Dans le sol, les champignons interagissent dynamiquement avec une multitude de microbes tels que d’autres champignons5, bactéries6, virus7 et nématodes8. Ces interactions ont des conséquences importantes sur la santé des sols et des plantes. Pourtant, en raison d’un manque de systèmes expérimentaux appropriés capables d’imager les micro-organismes en interaction avec une haute résolution, beaucoup restent indéfinis.
Les recherches concernant les interactions bactériens-fongiques (IFI) et les interactions fongiques-fongiques (IFF) ont des applications précieuses dans divers domaines, y compris les antimicrobiens en médecine et les agents de lutte biologique en agriculture. Par exemple, le champignon Coprinopsis cinerea produit le peptide copsine, dont il a été démontré qu’il présente une activité antibactérienne contre l’agent pathogène humain Listeria monocytogenes9. De même, le composé dérivé du champignon, la griséofulvine, est largement utilisé comme traitement des infections fongiques humaines et est en outre capable d’inhiber la croissance du champignon phytopathogène Alternaria solani10,11. Il a également été démontré que plusieurs souches de la bactérie bacillus subtilis sont des agents de lutte biologique efficaces de l’agent pathogène fongique des plantes Rhizoctonia solani12,13. Néanmoins, en raison des limites associées aux méthodologies traditionnelles, les IFI et les IFF sont mal compris au niveau des cellules individuelles.
Les études conventionnelles explorent généralement les IFI et les IFF à l’échelle macro en utilisant des plaques de gélose avec deux espèces ou plus en confrontation. Leur interaction est évaluée en mesurant les taux de croissance et la production de métabolites de l’espèce confrontante 14,15,16; cependant, cette méthodologie n’est résolue qu’au niveau de la colonie. Pour étudier les interactions au niveau cellulaire, les inoculants bactériens et fongiques peuvent être cultivés sur des lames de microscope en verre recouvertes d’agar qui sont ensuite imagées au microscope17. Néanmoins, il peut être difficile de suivre un seul hyphe à l’aide de lames de microscope en raison d’un manque de confinement, ce qui signifie que les images time-lapse sont plus difficiles à obtenir. En outre, la possibilité de confiner spatialement d’autres micro-organismes dans des régions définies du mycélium fongique ou de créer des environnements chimiques définis qui peuvent être perturbés, par exemple, n’est pas possible dans de telles configurations. La nature « boîte noire » du sol ajoute également à la complexité de l’étude des interactions fongiques-microbiennes au niveau des cellules individuelles18. En observant les espèces en interaction loin de l’incroyable diversité du microbiome du sol, la manière exacte dont les membres individuels interagissent peut être évaluée. Ainsi, il existe un besoin continu de plates-formes polyvalentes qui permettent l’imagerie unicellulaire à haute résolution des IFI et des IFF.
Les technologies microfluidiques, appelées systèmes de laboratoire sur puce, constituent une plate-forme idéale pour l’étude des BFI et des IFF au niveau des cellules individuelles. Le domaine de la microfluidique, issu des technologies développées pour l’analyse chimique et la microélectronique, a été adopté par les sciences biologiques19. Les technologies microfluidiques régulent de petits volumes de fluides au sein d’un réseau sur mesure de canaux miniaturisés, ayant au moins une dimension à l’échelle micrométrique, et leur utilisation dans la recherche biologique s’étend20. En particulier, des dispositifs microfluidiques ont été développés pour examiner la croissance des champignons filamenteux 21,22,23,24,25,26,27,28,29,30. L’un des avantages de l’utilisation de cette technologie est que le confinement des hyphes et la distribution des nutriments dans les microcanaux ressemblent davantage à la structure de l’environnement du sol que les méthodes conventionnelles d’agar31. Récemment, des plateformes microfluidiques ont été utilisées pour étudier les interactions entre les neutrophiles humains et les agents pathogènes fongiques32, les bactéries et les racines des plantes33, ainsi que les champignons et les nématodes34,35.
L’un des nombreux avantages de l’utilisation de la microfluidique pour étudier les interactions microbiennes comprend le contrôle spécifique de l’environnement des microcanaux. Par exemple, les régimes d’écoulement laminaire peuvent être exploités pour générer des gradients de concentration définis, ce qui est particulièrement utile lors de l’examen de la chimiotaxie bactérienne36. Un autre avantage est que la nature transparente du poly(diméthylsiloxane) (PDMS), un polymère élastomère biocompatible peu coûteux couramment utilisé dans la fabrication de dispositifs microfluidiques, facilite l’imagerie à haute résolution de cellules individuelles à l’aide de la microscopie à fond clair et à fluorescence37. De même, le confinement des microbes dans des microcanaux signifie que des expériences en accéléré de suivi de cellules individuelles peuvent être effectuées, permettant d’enregistrer et de quantifier les réponses cellulaires individuelles37. Enfin, comme les dispositifs microfluidiques peuvent être conçus pour être conviviaux, ils peuvent être facilement utilisés par des non-experts38.
Il est important de mieux connaître les interactions entre les micro-organismes vivant dans le sol pour améliorer les pratiques de gestion durable des écosystèmes qui maintiennent la biodiversité et atténuer l’impact des changements climatiques sur les environnements terrestres39. Ainsi, le développement de nouveaux outils microfluidiques est fondamental pour élargir la compréhension des champignons et de leurs interactions au niveau cellulaire. Le protocole ici se concentrera sur deux dispositifs microfluidiques produits pour l’étude des BFI40 et41 tels que représentés à la figure 1.
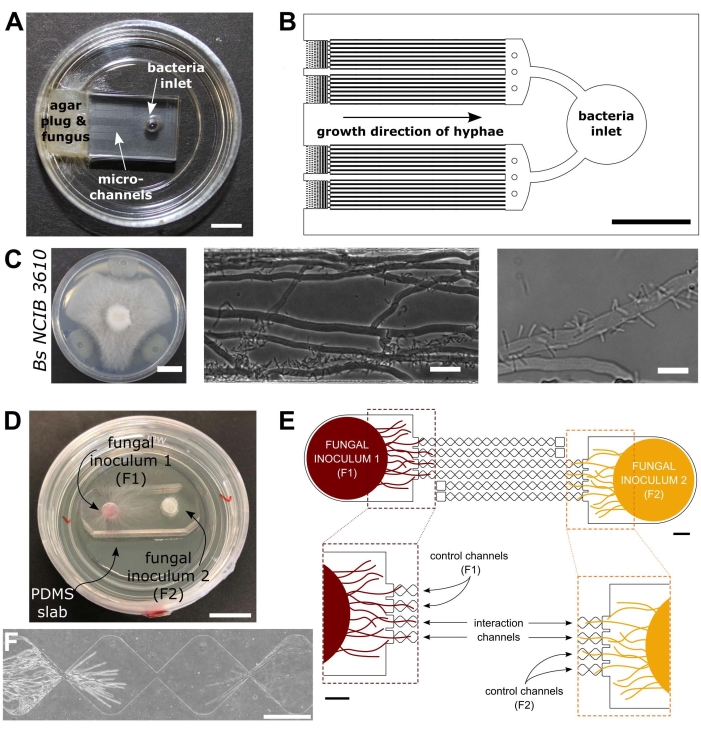
Figure 1 : Représentation visuelle et schématique des dispositifs d’interaction bactérienne-fongique (BFI) et d’interaction fongique (FFI). (A) Image du dispositif BFI. Un bouchon mycélien est placé à l’entrée d’une extrémité des microcanaux pour permettre la croissance hyphale dans l’appareil. L’entrée bactérienne se trouve à l’extrémité opposée. Barre d’échelle = 5 mm. (B) Vue d’ensemble schématique du dispositif BFI, décrivant le positionnement des entrées bactériennes et la direction de la croissance hyphale à travers les microcanaux d’interaction. Les canaux ont une profondeur de 10 μm, une largeur de 100 μm et une longueur de 7 mm, avec 28 canaux d’observation au total. (C) Essai de confrontation sur plaque de gélose entre Coprinopsis cinerea et Bacillus subtilis NCIB 3610, barre d’échelle = 20 mm (à gauche). Images microscopiques montrant l’interaction entre C. cinerea et B. subtilis NCIB 3610 dans le microcanal (milieu et droite), c’est-à-dire la fixation polaire des bactéries aux hyphes fongiques. Barre d’échelle = 25 μm (au milieu) et 10 μm (à droite). (D) Image du dispositif FFI collé à une boîte de Petri à fond de verre, double inoculée avec des bouchons mycéliens. Barre d’échelle = 1 cm. (E) Vue d’ensemble schématique du périphérique FFI. Deux bouchons d’inoculant fongiques sont introduits dans les entrées à chaque extrémité de l’appareil, permettant l’exploration hyphale des microcanaux. Les canaux de contrôle sont connectés à une seule entrée fongique et ont un canal sans issue, empêchant les interactions entre les champignons d’essai. Les canaux d’interaction relient les deux entrées fongiques et permettent des interactions hyphales entre les sujets testés au sein du microcanal. Chaque canal d’interaction se compose de 18 sections en forme de diamant, mesurant une longueur totale de 8,8 mm (490 x 430 μm par diamant), 10 μm de profondeur et ayant une région de connexion entre chaque diamant de 20 μm. Les types de canaux sont dupliqués, barres d’échelle = 1 mm. (F) Zone d’interaction entre deux fronts hyphales approchants, se développant à partir des extrémités opposées du canal d’interaction interconnecté. Image de microscopie à contraste de phase, barre d’échelle = 250 μm. Les panneaux de cette figure ont été modifiés à partir de Stanley et al., 2014 (A-C)40 et Gimeno et al., 2021 (D-F)41. Veuillez cliquer ici pour voir une version agrandie de cette figure.
