Leptospirose bij de mens is voornamelijk afkomstig van milieubronnen 1,2. De aanwezigheid van Leptospira in meren, rivieren en beken is een indicator van de overdracht van leptospirose onder dieren in het wild en huisdieren en productiedieren die uiteindelijk in contact kunnen komen met deze watermassa’s 1,3,4. Bovendien is Leptospira geïdentificeerd in niet-natuurlijke bronnen, waaronder rioolwater, stilstaand en kraanwater 5,6.
Leptospira is een wereldwijd verspreide bacterie 7,8, en de rol van het milieu bij het behoud en de overdracht ervan is algemeen erkend. Leptospira kan overleven in drinkwater met een variabele pH en mineralen9, en in natuurlijke waterlichamen1. Het kan ook lange tijd overleven in gedestilleerd water10, en onder een constante pH (7,8) kan het tot 152 dagen overleven11. Bovendien kan Leptospira interageren in bacteriële consortia om barre omstandigheden te overleven12,13. Het kan deel uitmaken van biofilms in zoet water met Azospirillum pt Sphingomonas en is zelfs in staat om te groeien en temperaturen van meer dan 49 °C14,15 te doorstaan. Het kan zich ook vermenigvuldigen in drassige grond en tot 379 dagen levensvatbaar blijven16, waardoor het vermogen om de ziekte te veroorzaken tot een jaar lang behouden blijft17,18. Er is echter weinig bekend over de ecologie in waterlichamen en hoe deze daarin is verdeeld.
Sinds de ontdekking was de studie van het geslacht Leptospira gebaseerd op serologische tests. Pas in de huidige eeuw kwamen moleculaire technieken steeds vaker voor in de studie van deze spirochaete. De dot-blot is nauwelijks gebruikt voor zijn identificatie met behulp van (1) een isotopische sonde op basis van het 16S rRNA en op een inter-simple sequence repeat (ISSR)19,20, (2) als een op nanogoud gebaseerde immunoassay voor menselijke leptospirose toegepast op urine21, of (3) als een op antilichamen gebaseerde test voor runderurinemonsters22. De techniek raakte in onbruik omdat deze oorspronkelijk gebaseerd was op isotopische sondes. Het is echter een bekende techniek die, in combinatie met PCR, verbeterde resultaten oplevert en als veilig wordt beschouwd vanwege het gebruik van niet-isotopische sondes. PCR speelt een cruciale rol bij de verrijking van het Leptospira-DNA door een specifiek DNA-fragment te amplificeren dat in sporenhoeveelheden in een monster kan worden aangetroffen. Tijdens elke PCR-cyclus wordt de hoeveelheid van het beoogde DNA-fragment in de reactie verdubbeld. Aan het einde van de reactie is het amplicon vermenigvuldigd met een factor van meer dan een miljoen23. Het door PCR geamplificeerde product, vaak niet zichtbaar bij agarose-elektroforese, wordt zichtbaar door specifieke hybridisatie met een DIG-gelabelde sonde in de dot-blot 24,25,26.
De dot-blot-techniek is eenvoudig, robuust en geschikt voor tal van monsters, waardoor het toegankelijk is voor laboratoria met beperkte middelen. Het is gebruikt in een verscheidenheid aan bacteriestudies, waaronder (1) orale bacteriën27, (2) andere soorten monsters zoals voedsel en ontlasting28, en (3) de identificatie van niet-kweekbare bacteriën29, vaak in overeenstemming met andere moleculaire technieken. Een van de voordelen van de dot-blot-techniek zijn: (1) Het membraan heeft een hoge bindingscapaciteit, in staat om meer dan 200 μg/cm2 nucleïnezuren en tot 400 μg/cm2 te binden; (2) Dot-blot-resultaten kunnen visueel worden geïnterpreteerd zonder dat er speciale apparatuur nodig is, en (3) ze kunnen gemakkelijk jarenlang bij kamertemperatuur (RT) worden bewaard.
Het geslacht Leptospira is ingedeeld in pathogene, intermediaire en saprofytische clades30,31. Het onderscheid tussen deze clades kan worden bereikt op basis van specifieke genen zoals lipL41, lipL32 en het 16S rRNA. LipL32 is aanwezig in de pathogene clades en vertoont een hoge gevoeligheid in verschillende serologische en moleculaire instrumenten, terwijl het afwezig is in saprofytensoorten21. Het huishoudgen lipL41 staat bekend om zijn stabiele expressie en wordt gebruikt in moleculaire technieken32, terwijl het 16S rRNA-gen wordt gebruikt voor hun classificatie.
Deze methodologie kan worden toegepast op grote hoeveelheden water nadat ze zijn geconcentreerd door centrifugeren. Het maakt het mogelijk om verschillende punten en diepten in een waterlichaam te beoordelen om de aanwezigheid van leptospiraalvormig DNA en de clade waartoe het behoort te detecteren. Deze tool is waardevol voor zowel ecologische als algemene screeningsdoeleinden en kan ook worden gebruikt om andere niet-kweekbare bacteriën te detecteren die in water aanwezig kunnen zijn.
Bovendien zijn PCR- en dot-blot-assays technisch en economisch betaalbaar voor een breed scala aan laboratoria, zelfs voor laboratoria die niet over geavanceerde of dure apparatuur beschikken. Deze studie heeft tot doel de op digoxigenine gebaseerde dot-blot toe te passen voor de identificatie van de drie Leptospira-clades in watermonsters die zijn verzameld uit natuurlijke waterlichamen.
Bacteriestammen
Twaalf Leptospira serovars (Autumnalis, Bataviae, Bratislava, Canicola, Celledoni, Grippothyphosa, Hardjoprajitno, Icterohaemorrhagiae, Pomona, Pyrogenes, Tarassovi en Wolffi) werden in deze studie opgenomen. Deze serovars maken deel uit van de collectie van de afdeling Microbiologie en Immunologie, Faculteit Diergeneeskunde en Zoötechniek, Nationale Autonome Universiteit van Mexico, en ze worden momenteel gebruikt in de microagglutinatietest (MAT).
Alle Leptospira-serovars werden gekweekt in EMJH en hun DNA werd geëxtraheerd met behulp van een commerciële DNA-extractiekit (zie Materiaaltabel). Een genomische DNA-mix van de twaalf serovars werd gebruikt als positieve controle voor de pathogene clade van Leptospira . Als positieve controle van de Leptospira-intermediaire clade werd ook genoom-DNA van Leptospira fainei serovar Hurstbridge-stam BUT6 opgenomen, en als positieve controle voor de Leptospira-saprofytenclade werd ook genomisch DNA van Leptospira biflexa serovar Patoc-stam Patoc I opgenomen.
Negatieve controles bestonden uit een leeg plasmide, DNA van niet-verwante bacteriën (Ureaplasma urealyticum, Staphylococcus aureus, Brucella abortus, Salmonella typhimurium, Shigella boydii, Klebsiella pneumoniae, Acinetobacter baumannii pt Escherichia coli), en water van PCR-kwaliteit, dat diende als niet-sjablooncontrole.
Monsters van water
Twaalf proefmonsters werden verzameld met behulp van een gestratificeerde-lukrake bemonsteringsmethode van het Cuemanco Biological and Aquaculture Research Center (CIBAC) (19° 16′ 54″ N 99° 6′ 11″ W). Deze monsters werden verkregen op drie diepten: oppervlakkig, 10 en 30 cm (figuur 1A, B). De procedures voor het verzamelen van water hadden geen invloed op bedreigde of beschermde diersoorten. Elk monster werd verzameld in een steriele microcentrifugebuis van 15 ml. Om het monster te verzamelen, werd elke buis voorzichtig ondergedompeld in het water, op de geselecteerde diepte gevuld en vervolgens verzegeld. De monsters werden op 22 °C gehouden en onmiddellijk naar het laboratorium vervoerd voor verwerking.
Elk monster werd geconcentreerd door centrifugatie in steriele microcentrifugebuisjes van 1,5 ml bij 8000 x g gedurende 20 minuten bij kamertemperatuur. Deze stap werd herhaald totdat alle monsters waren geconcentreerd in één buis, die vervolgens werd gebruikt voor DNA-extractie (Figuur 1C).
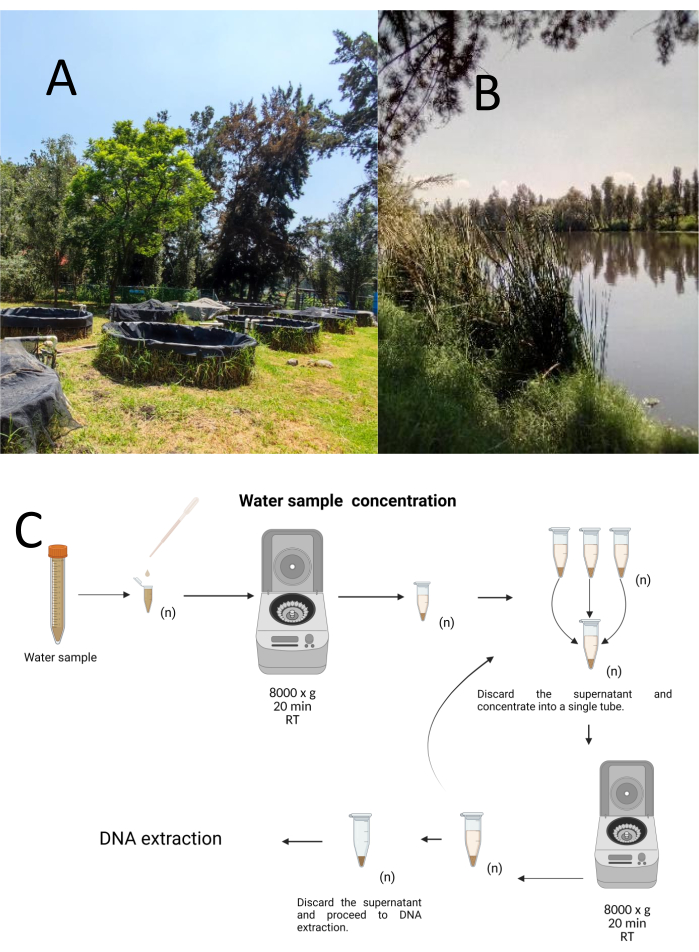
Figuur 1: Concentratie van watermonsters door centrifugatie. (A) Waterbemonsteringsvijvers en (B) Natuurlijke beken. (C) Verwerking van watermonsters op basis van centrifugatie in herhaalde stappen, zo vaak als nodig is (n). Klik hier om een grotere versie van deze figuur te bekijken.
DNA-extractie
Totaal DNA werd geïsoleerd met behulp van een in de handel verkrijgbare genomische DNA-kit volgens de instructies van de fabrikant (zie Materiaaltabel). DNA-extracties werden geëlueerd in 20 μL elutiebuffer en de DNA-concentratie werd bepaald door een UV-spectrofotometer bij 260-280 nm en bewaard bij 4 °C tot gebruik.
PCR-amplificatie
De PCR-doelen waren de 16S rRNA-, lipL41- en lipL32-genen, die DNA van het geslacht Leptospira identificeren en het onderscheid tussen de drie clades mogelijk maken: pathogeen, saprofytische en intermediairen. Zowel de primers als de sondeontwerpen waren gebaseerd op de eerdere werken van Ahmed et al., Azali et al., Bourhy et al., Weiss et al., en Branger et al.33,34,35,36,37. De volgorde van elke sonde, primer en geamplificeerd fragment wordt beschreven in tabel 1 en hun uitlijning met referentiesequenties wordt gegeven in aanvullend bestand 1, aanvullend bestand 2, aanvullend bestand 3, aanvullend bestand 4 pt aanvullend bestand 5. De PCR-reagentia en thermocyclusvoorwaarden worden beschreven in het protocolgedeelte.
Amplificatieproducten werden gevisualiseerd door elektroforetische scheiding op een 1% agarosegel in TAE (40 mM Tris-base, 20 mM azijnzuur en 1 mM EDTA; pH 8,3), bij 60 V gedurende 45 minuten met ethidiumbromidedetectie, zoals weergegeven in aanvullende figuur 1. Genoom-DNA verkregen van elke serovar werd gebruikt met concentraties variërend van 6 x 106 tot 1 x 104 genomische equivalente kopieën (GEq) in elke PCR-reactie, gebaseerd op de genoomgrootte van L. interrogans (4, 691, 184 bp)38 voor pathogene Leptospira, de genoomgrootte van L. biflexa (3, 956, 088 bp)39 voor saprofytische Leptospira, en de genoomgrootte van L. fainei serovar Hurstbridge-stam BUT6 (4, 267, 324 bp) met toetredingsnummer AKWZ00000000.2.
De gevoeligheid van de sondes werd beoordeeld met DNA van elke pathogene serovar, L. biflexa serovar Patoc stam Patoc I en L. fainei serovar Hurstbridge-stam BUT6 in elk experiment. Om de specificiteit van de PCR- en dot-blot-hybridisatietest te beoordelen, werd DNA van niet-verwante bacteriën opgenomen.
Tabel 1: PCR-primers en -sondes om producten te amplificeren voor het identificeren van de pathogene, saprofyt- en intermediaire clades van Leptospira. Klik hier om deze tabel te downloaden.
Dot-blot hybridisatie-test
De techniek wordt dot-blot genoemd omdat de gaten waarin het DNA-monster wordt geplaatst een puntvorm hebben, en wanneer ze worden opgezogen om op hun plaats te worden gefixeerd door vacuümzuiging, krijgen ze deze vorm. Deze techniek is ontwikkeld door Kafatos et al.40. De techniek maakt de semi-kwantificering van Leptospira in elk PCR-positief monster mogelijk. Het protocol bestaat uit een denaturatie met NaOH 0,4 M bij kamertemperatuur, monsters met Leptospira-DNA van 30 ng tot 0,05 ng, overeenkomend met 6 x 106 tot 1 x 104 leptospiren, worden gedept op een nylon membraan met een dot-blot-apparaat met 96 putjes. Na immobilisatie wordt het DNA aan het membraan gebonden door blootstelling aan 120 mJ UV-licht. Elke DNA-sonde wordt geconjugeerd met digoxigenine-11 dUTP door een terminale transferasekatalysestap aan het 3′-uiteinde (Digoxigenine is een plantaardige steroïde verkregen uit Digitalis purpurea, gebruikt als verslaggever41). Na de strikte hybridisatie van de gelabelde DNA-sonde (50 pmol) bij de specifieke temperatuur op het doel-DNA, worden de DNA-hybriden gevisualiseerd door de chemiluminescentiereactie met het anti-digoxigenine alkalische fosfatase-antilichaam dat covalent is geconjugeerd met zijn substraat CSPD. De luminescentie wordt vastgelegd door blootstelling aan een röntgenfilm (figuur 2).
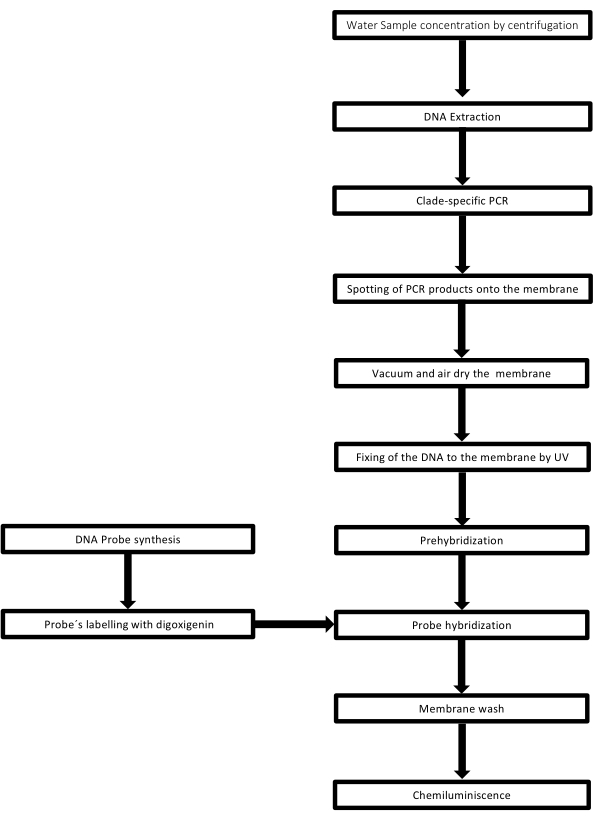
Figuur 2: Stappen van de procedure voor de PCR-dot-blot-test. Klik hier om een grotere versie van deze figuur te bekijken.
