Celler inneholder et mangfold av biomacromolecules som hele tiden kommuniserer med hverandre. Denne assosiasjonen gir opphav til komplekser som deltar i de cellulære veier som er ansvarlige for deres funksjon i signaltransduksjon, regulering av genekspresjon og cellevandring blant andre. Alt protein-protein interaksjoner som forekommer i en celle omfatter et nettverk kjent som interactome. I Saccharomyces cerevisiae mer enn 70% av sitt protein har vist seg å ha samvirkende partnerne 1. Forstå interactome av en celle og deres funksjoner gi relevant informasjon om kompleksiteten og mangfoldet av levende organismer. Flere metoder har blitt beskrevet å identifisere og karakterisere protein-protein interaksjoner. Forskjellig høy gjennom benyttet metoder som gjær to-hybrid 2, protein-fragment komplemente assays 3, affinitetsrensing 4 koblet til massespektrometri og protein microarrays brukes til å identifisere en interaksjon 5,6. Når identifisert, er det nødvendig å validere det, og dette kan variere fra sak til sak. Vanligvis disse eksperimentene involverer forstyrre samspillet selv på nivå med de enkelte medlemmer av samspillet paret, for eksempel ved genet sletting eller overekspresjon av en av proteiner, og deretter se etter endringer i egenskaper eller funksjon av den andre medlemmet på cellenivå. Deretter blir biofysiske teknikker 7 som brukes til å karakterisere protein-protein interaksjon på molekylært nivå. For dette formål, er strukturen av proteinkomplekser bestemt ved røntgen-krystallografi, kjernemagnetisk resonans og kryo-elektronmikroskopi mens kalorimetri og fluorescens-spektroskopi blir brukt for å kvantitativt og mekanistisk beskrive dem.
I dette arbeidet ble fluorescens anisotropi brukt som en teknikk for å karakterisere samspillet mellom GTPase EFL1 og SBDS-proteinet. Disse proteinene deltar i syntesen av ribosomer ved å fremme frigivelse av eukaryotisk initiering faktor 6 fra overflaten av den 60S ribosomale subenhet 8. Den SBDS proteinet er mutert på en sykdom som er kjent som den Shwachman-Diamond syndrom 9 og virker som et guanin nukleotid utveksling faktor for EFL1 avtagende dets affinitet for guanosin difosfat 10,11. Sykdoms mutasjoner i SBDS avskaffe samspillet med EFL1 og dermed hindre aktivering.
Fluorescens anisotropi er ofte brukt i biologiske anvendelser for å studere protein-peptid eller protein-nukleinsyre-interaksjoner. Den er basert på det prinsipp at en fluoroforen eksiteres med polarisert lys resulterer i en delvis polarisert utslipp. Fluorescens anisotropi er definert ved ligning 1:
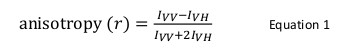
der jeg vv og jeg VH erfluorescensstyrkene den vertikalt (VV) og horisontalt (VH) polarisert utslipp når prøven er spent med vertikalt polarisert lys 12. Fluorescens anisotropi er følsom for faktorer som påvirker frekvensen av rotasjons diffusjon av fluoroforen og således avhenger av temperaturen, viskositeten av løsningen og den tilsynelatende molekylstørrelse av fluoroforen. Den tilsynelatende størrelse av et protein inneholdende en fluorofor øker når den kommuniserer med et annet protein og en slik endring kan deretter evalueres som en endring i anisotropi. Nærmere bestemt vil en fluorofor som roterer langsomt i oppløsning i forhold til dets fluoriserende levetid vil ha en stor I VV verdi og liten jeg VH verdi, og derfor vil oppvise en forholdsvis stor anisotropi. For fluorophores som tørkes raskt i forhold til deres fluorescerende levetid, jeg vv og jeg VH vil være like og deres anisotropi verdien vil være liten 12 </sup> (Figur 1). I tillegg, for en god anisotropi signal til støymåling, er det nødvendig å ha en fluorofor med en fluorescens levetid lik den rotasjonskorrelasjonstiden til molekylet av interesse. Ellers er det ikke mulig å nøyaktig registrere forskjellen i anisotrofien mellom den frie protein, og at i komplekset. For eksempel, anisotropien av en fluorescerende probe med en levetid nær 4 nsek som for eksempel fluorescein eller rhodamin festet til en lavmolekylær forbindelse med 100 Da er 0,05. Binding til et molekyl av 160 kDa vil øke dens anisotropi verdien til 0,29; en forskjell som kan måles nøyaktig. I motsetning til dette, vil det samme fluorescerende probe som er involvert i en bindingsreaksjon, som plutselig økning i molekylstørrelse varierer fra 65 til 1000 kDa bare resultere i en endring anisotropi på 0,28 til 0,3, noe som er for små til å måles nøyaktig. I dette scenariet vil en sonde med en levetid på 400 ns være mer egnet 12.
<pclass = "jove_content">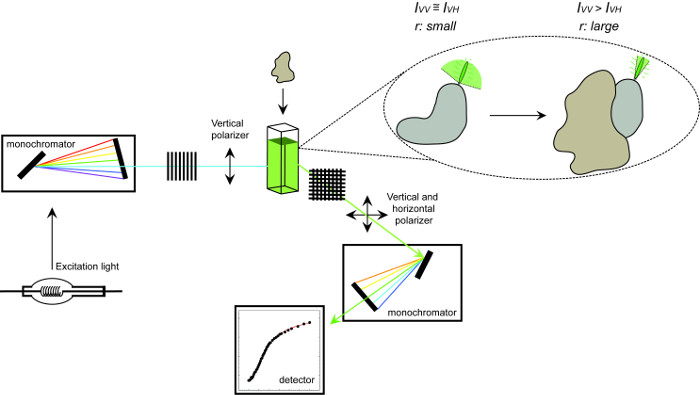
Figur 1. Skjematisk representasjon av utstyret som brukes til å måle fluorescens anisotropi og prosedyren. Skjematisk representasjon av utstyret som brukes til å utføre en protein-protein interaksjon eksperiment som måler fluorescensen anisotropi. Fluoroforer som ristes rask visnings liten anisotropi som øker ved binding til et samspill partner. Klikk her for å se en større versjon av dette tallet.
Fluorescensanvendelser krever tilstedeværelse av en fluorofor i hvilket som helst av molekylene studert. For å undersøke protein-protein interaksjoner er det tre typer fluoroforer: 1) tryptofanrester som er tilstede i proteiner, 2) kjemisk festet fluoroforer og 3) fluoriserende fusjonspartnere som grønt fluorescerende protein (GFP) og dens derivatanter. De fleste proteiner har tryptofanrester på sin struktur, og dermed den enkleste måten for å måle en interaksjon er ved å overvåke endringene i den tilsvarende fluorescensspektra eller ved å overvåke endringer i fluorescens-intensiteten av tryptofanrester. Imidlertid kan tryptofanrester være til stede i begge proteiner kompliserer analysen. På den annen side, for en fluorofor til å endre sin fluoriserende egenskaper på grunn av en interaksjon det er behov for å være plassert på eller nær bindingssetet og det kunne interferere med interaksjonen selv. Dette trenger spesiell oppmerksomhet når du bruker store fluoroforer som GFP. Hvis ingen av disse fluoroforer kan brukes til bindingsstudier er det nødvendig, da, å innføre ytre fluoroforer til den ene av de involverte proteiner. Mange kjemisk syntetiserte fluoroforer eksistere og kan være kovalent bundet til proteiner via sine reaktive grupper slik som aminogrupper (sidekjede av lysin eller N-terminus) og tiol- grupper i cystein. Fluorophore derivater med isothiocyanate og suksinimidyl estere reagere med amidgrupper mens iodoacetamide og maleimide er tiol-reaktive grupper 13. De mest vanlige fargestoffer som brukes i fluorescensanvendelser er derivater av fluorescein og rhodamin grønne farvestoffer, koumariner, BODIPY fluoroforer og Alexa Fluor fargestoffer. En detaljert liste over kommersielt tilgjengelige fluoroforer og deres anvendelse kan finnes i referanser 14,15. For vellykket merking, må den reaktive gruppe være eksponert på overflaten av proteinet, men på grunn av det store antall reaktive funksjonelle grupper som typisk er tilstede i polypeptidene det er svært vanskelig å få sete-spesifikk modifisering. Proteinet av interesse i denne studien, SBDS, inneholder 5 gratis cysteiner og 33 lysines som kan resultere i flere språk merking. Ikke-uniform merking kan påvirke binding og vil komplisere dataanalyse som ulike fluoroformolekyler kan lokke fram ulike fluorescerende intensitet signaler ved binding. Overcome dette problemet, brukte vi Flash fluorophore, 4 ', 5'-bis (1,3,2 dithioarsolan-2-yl) fluorescein til nettstedet Direkt label SBDS protein. Dette er en arsenoxide fargestoff med en høy affinitet for fire adskilte cysteiner i et motiv kjent som flash-kode som består av sekvensen CCXXCC hvor X er hvilken som helst aminosyre annen enn cystein 16,17. Dette tetracysteine motiv tilsettes til N- eller C-terminale enden av proteinet ved hjelp av genteknologi sammen med en egnet linker for å forhindre forstyrrelse av den samlede fold av proteinet. Paret består av Flash fargestoff og Flash-koden ble opprinnelig utviklet for å stedsspesifikke label proteiner i levende celler 17, men det kan også brukes til å merke rensede proteiner in vitro som det er eksemplifisert her. I tillegg har enzymatiske strategier også blitt utviklet for å muliggjøre stedsspesifikke funksjonalisering av proteiner 18.
I dette manuskriptet beskriver vi nytten av fluorescens anisotropi enet verktøy for å studere protein-protein interaksjoner. Binding kan vurderes ved enkel inspeksjon av bindingen kurveformen mens kvantitativ informasjon kan fås fra tilpasning av de eksperimentelle data.
