Muligheten til å ingeniør genomet av enhver levende organisme har mange biomedisinsk og bioteknologisk programmer, for eksempel korrigering av sykdomsfremkallende mutasjoner, bygging av nøyaktige mobilnettet modeller for sykdom studier eller generering av landbruket vekster med ønskelig egenskaper. Siden begynnelsen av århundret, ulike teknologier er utviklet for genomet engineering i pattedyrceller, inkludert meganucleases1,2,3, sink finger nucleases4,5, eller transkripsjon aktivator som effektor nucleases (TALENs)6,7,8,9. Men er disse tidligere teknologiene vanskelig å programmet eller kjedelig å montere, og dermed hindrer deres utbredt adopsjon i forskning og industri.
De siste årene, de klynget interspaced regelmessig kort palindromic gjentar (CRISPR) – CRISPR-assosiert (Cas) systemet har dukket opp som en kraftig ny genomet engineering teknologi10,11. Opprinnelig en adaptive immunsystemet i bakterier, det har blitt deployert for genomet endring i planter og dyr, inkludert mennesker. Hovedårsak hvorfor CRISPR-Cas har fått så mye popularitet i så kort tid er at elementet som bringer den viktigste Cas-endonuclease, som Cas9 eller Cas12a (også kjent som Cpf1), til riktig sted i genomet er bare et kort stykke chimeric enkelt støttelinje RN En (sgRNA), som er enkelt å utforme og billig å syntetisere. Etter blir rekruttert til målområdet, Cas enzymet fungerer som et par molekylær saks og innstiftet bundet DNA med sin RuvC, HNH eller Nuc domener12,13,14. Resulterende dobbel strandet pause (DSB) repareres senere av cellene via enten ikke-homologe slutten med (NHEJ) eller homologi-rettet reparasjon (HDR) veien. I fravær av en reparasjon mal repareres DSB av feilutsatte NHEJ veien, som kan gi opphav til pseudo-tilfeldige innsetting eller sletting av nukleotider (indeler) på webområdet kutt, potensielt forårsake frameshift mutasjoner i protein-koding gener. Men i nærvær av en donor mal som inneholder endringene i DNA, er DSB reparert av Hi-Fi HDR veien. Vanlige typer donor maler inkluderer enkelt-strandet oligonucleotides (ssODNs) og plasmider. Tidligere brukes vanligvis hvis tiltenkte DNA endringene er små (for eksempel endring av en enkelt base-par), mens sistnevnte brukes vanligvis hvis man ønsker å sette inn en relativt lang sekvens (for eksempel koding sekvensen av en grønn fluorescerende protein eller GFP) i målet locus.
Endonuclease aktiviteten til Cas protein krever tilstedeværelse av en protospacer tilstøtende motiv (PAM) mål område15. PAM Cas9 er på 3 slutten av protospacer, PAM Cas12a (også kalt Cpf1) er på 5′ slutten i stedet16. Cas-guiden RNA komplekse er ikke innføre en DSB hvis PAM er fraværende17. Derfor plasserer PAM en begrensning på genomisk stedene der en bestemt Cas-nuclease er i stand til å holde seg. Heldigvis viser Cas nucleases fra ulike bakterie-art vanligvis ulike PAM krav. Ved å integrere ulike CRISPR-Cas systemer i våre tekniske verktøykassen, kan vi derfor utvide omfanget av nettsteder som kan målrettes i et genom. Videre kan et naturlig Cas enzym være konstruert eller utviklet for å gjenkjenne alternativ PAM sekvenser, ytterligere utvide omfanget av genomisk mål tilgjengelig for manipulasjon18,19,20.
Selv om flere CRISPR-Cas-systemer finnes genomet engineering formål, har de fleste brukere av teknologi stolt på Cas9 nuclease fra Streptococcus pyogenes (SpCas9) av flere grunner. Først krever det en relativt enkelt NGG PAM, i motsetning til mange andre sertifiseringsinstanser proteiner som kan bare holde seg i nærvær av mer komplekse PAMs. Det andre er den første Cas endonuclease å bli distribuert i menneskeceller21,22,23,24. Tredje er SpCas9 langt den beste preget enzymet hittil. Hvis en forsker ønsker å bruke en annen Cas nuclease, vil han eller hun ofte være uklart om hvordan best å utforme eksperimentet og hvor godt andre enzymer vil opptre i ulike biologiske sammenhenger i forhold til SpCas9.
For å utdype den relative ytelsen til forskjellige CRISPR-Cas-systemer, har vi nylig utførte en systematisk Sammenlikning av fem Cas endonucleases-SpCas9, Cas9 enzymet fra Staphylococcus aureus (SaCas9), Cas9 enzymet fra Neisseria meningitidis (NmCas9), Cas12a enzymet fra Acidaminococcus sp. BV3L6 (AsCas12a) og Cas12a enzymet fra Lachnospiraceae bakterie ND2006 (LbCas12a)25. For en rettferdig sammenligning vurdert vi de ulike Cas nucleases bruker samme sett med målområder og andre eksperimentelle forhold. Studien også avgrenset design parameterne for hver CRISPR-Cas-system, som ville tjene som en nyttig referanse for brukere av teknologi. Her, bedre aktiverer forskere å bruke CRISPR-Cas system, gir vi en trinnvis protokoll for optimal genomet engineering med forskjellige Cas9 og Cas12a enzymer (se figur 1). Protokollen inkluderer ikke bare eksperimentelle detaljer men også viktige Utformingshensyn å maksimere sannsynligheten for et vellykket genomet engineering resultat i pattedyrceller.
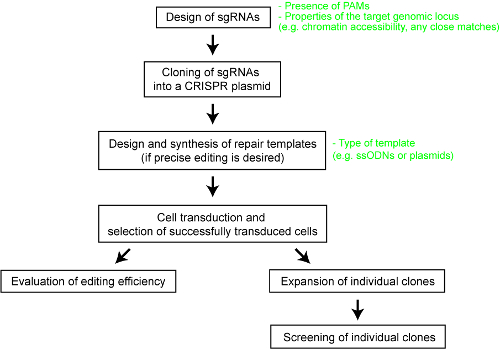
Figur 1 : En oversikt over arbeidsflyten skal generere genomet endret humane cellelinjer. Klikk her for å se en større versjon av dette tallet.
