由于超过 80% 的病例具有压倒性的散发性,再加上已知是致病性的大量基因突变2,因此在实验室中对肌萎缩性侧索硬化症 (ALS) 进行建模极具挑战性。尽管如此,所有ALS病例都有一个统一的特征,即在完全神经元变性之前,突触前运动神经元和突触后肌肉细胞之间存在功能障碍的通信3,4。临床上,当患者失去剩余的上下运动神经元的连接时,它们在整个疾病过程中表现出神经元过度和低兴奋性的特征5,6,7,8,9,反映了这些突触的复杂潜在分子变化,我们作为ALS研究人员试图理解这一点。
多种转基因模型表明,神经肌肉接头的恶化和紊乱随着ALS致病基因突变的表达而发生,包括SOD110,FUS11,12,C9orf7213,14,15,16和TDP4317,18,19 通过形态学评估,包括突触胸顿、脊柱密度和突触前/突触后组织的评估。从机制上讲,自20世纪30年代科尔,霍奇金和赫胥黎的里程碑式论文以来,还可以通过体外细胞培养或组织切片制剂中的电生理技术评估突触反应20。通过这些策略,许多ALS模型已经显示出突触传递缺陷。例如,TDP43的突变变体导致NSC-34(脊髓x神经母细胞瘤杂交细胞系34)运动神经元样细胞中的放电频率增强并降低动作电位阈值21。在小鼠模型中,这种相同的变异还会导致神经肌肉接头(NMJ)处的功能失调性突触传递,然后才出现行为运动缺陷22。先前显示,在运动缺陷发生之前,FUS-ALS果蝇模型中的突变FUS表达导致NMJ突触传递减少11。最近一份使用来自C9orf72扩增载体的诱导多能干细胞的报告显示,易释放的突触囊泡池减少23。总而言之,这些研究和其他研究强调了对ALS疾病相关模型中突触信号传导的潜在机制建立更全面理解的重要性。这对于了解ALS的病理生物学和为患者开发潜在的治疗靶点至关重要。
电流和电压钳位单元的方法在确定膜性质方面具有无可估量的价值,例如电导率,静息膜电位和单个突触的量子含量20,24。然而,电生理学的一个重大局限性是,它在技术上具有挑战性,并且一次只能提供来自单个神经元的见解。活细胞共聚焦显微镜,加上特定的荧光探针,为以时空方式研究神经元的突触传递提供了机会25,26,27。虽然不是神经元兴奋性的直接测量,但这种荧光方法可以相对测量突触功能的两种分子相关性:突触囊泡释放和突触末端的钙瞬变。
当动作电位到达神经元的突触前末端区域时,钙瞬变被触发,促进从电信号到神经递质释放过程的转变28。定位于这些区域的电压门控钙通道严格调节钙信号传导,以调节神经递质释放的动力学29。首次报道的基于荧光的钙瞬变记录是使用双波长指示剂Fura-2 AM或单波长染料Flu-3 AM30,31,32进行的。虽然这些染料在当时提供了很好的新见解,但它们存在一些局限性,例如细胞内非特异性区室化,标记细胞的主动或被动染料损失,光漂白以及长时间成像的毒性33。在过去的十年中,遗传编码的钙指示剂已成为对各种形式的神经元活动进行成像的主力军。这些指示剂将修饰的荧光蛋白与钙螯合蛋白结合,钙螯合蛋白在Ca2 + 离子结合后快速切换荧光强度34。这些新指标的应用范围很广,可以在体外和体内环境中更容易地可视化细胞内钙瞬变。这些基因编码报告基因的一个家族,称为GCaMP,现在被广泛使用。这些指示剂包含C端钙调蛋白结构域,后跟绿色荧光蛋白(GFP),并被N端钙调蛋白结合区覆盖35,36。钙与钙调蛋白结构域的结合触发与钙调蛋白结合区域的相互作用,导致整体蛋白质结构的构象变化和GFP部分荧光的显着增加35,36。多年来,这个报告员家族经历了几次演变,以便为具有特定动力学(慢,中和快)的特定钙瞬变提供不同的读数,每个都具有略微不同的性质37,38。在这里,已经强调了报告者GcaMP6的使用,其先前已被证明可以在体内和体外检测神经元中的单次动作电位和树突状钙瞬变37。
突触前区域的钙瞬变触发突触囊泡融合事件,导致神经递质释放到突触中并在突触后细胞中启动信号传导事件28,39。突触囊泡既能快速释放又可循环,因为细胞稳态地维持稳定的细胞膜表面积和易于释放的融合池,具有膜结合的囊泡40。这里使用的苯乙烯基染料对脂质膜具有亲和力,并根据周围脂质环境的顺序特异性地改变其发射特性41,42。因此,它是标记回收突触囊泡并随后跟踪这些囊泡的理想工具,因为它们随后在神经元刺激后释放41,42。已经生成和优化的方案是对Gaffield及其同事最初描述的概念的改编,这使我们能够随着时间的推移连续地可视化苯乙烯染料标记的突触囊泡点41。
在这里,描述了两种相关的基于荧光的方法,可靠地报告了突触传递中涉及的特定细胞事件。已经定义了方案来探测培养神经元中去极化介导的突触前终末期钙流入和突触囊泡胞吐作用的动力学。在这里,方法和代表性结果侧重于使用原代啮齿动物皮质或运动神经元作为体外模型系统,因为有已发表的使用这些细胞类型的研究43,44。然而,这些方法也适用于分化的人类i3皮质样神经元45,因为我们在实验室目前正在进行的实验中也成功地使用了这两种方案。一般协议以逐步线性格式概述,如图1所示。简而言之,为了研究神经突起中的钙动力学,成熟的神经元被质粒DNA转染,以在巨细胞病毒(CMV)启动子下表达荧光报告GCaMP6m37,46。转染的细胞具有低水平的基底绿色荧光,在钙的存在下增加。指定感兴趣的区域以监测整个操作过程中的荧光变化。这允许测量钙的高度空间和时间局部波动37,46。为了评估突触囊泡融合和释放,成熟的神经元被加载到掺入突触囊泡膜中的苯乙烯染料,因为它们被回收,重整并重新加载突触前细胞中的神经递质41,42,43,47,48。目前用于此目的的染料沿着神经突触囊泡标记,并在实时成像实验中用作这些区域的代表,正如Kraszewski及其同事对苯乙烯染料和突触标记素的共染色所显示的那样49。这里包括也进行了类似染色的代表性图像(图2A)。以前的研究人员广泛使用这种染料来报告神经肌肉接头和海马神经元的突触囊泡动力学48,49,50,51,52,53,54,55,56.通过选择含染料囊泡的点状区域并监测囊泡释放后荧光强度的降低,可以研究刺激后的功能突触传递能力和释放的时间动力学43。对于这两种方法,使用含有高浓度氯化钾的培养基来去极化细胞以模仿神经元活动。指定成像参数以捕获跨越基线归一化和刺激捕获周期的亚秒间隔。确定每个时间点的荧光测量值,归一化到背景,并在实验时间段内进行量化。钙流入介导的GCaMP6m荧光增加或有效的突触囊泡胞吐作用苯乙烯染料释放荧光减少可以通过该策略检测到。下面描述了这两种方案的详细方法设置和参数,并讨论了它们的优点和局限性。
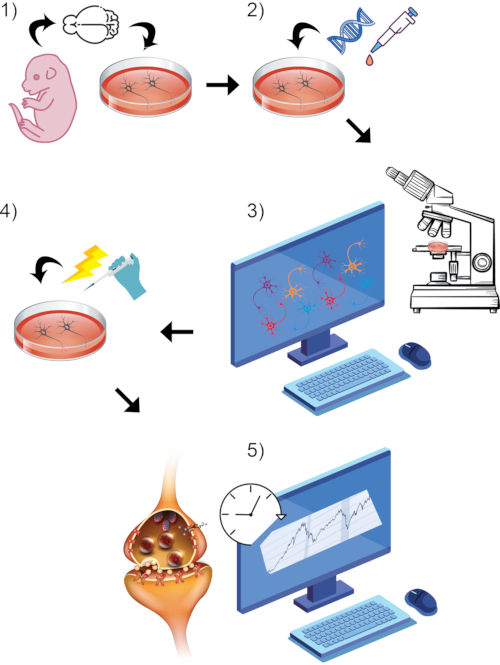
图 1:整体通用协议流程的可视化呈现。 (1) 在体外 分离并培养原代啮齿动物神经元到选定的成熟时间点。(2)引入GCaMP DNA或苯乙烯基染料作为突触活性的报告者。(3)使用配备实时成像的共聚焦显微镜和相关软件设置成像范例。开始基线记录期。(4)当细胞仍在进行实时图像捕获时,通过高KCl浴灌注刺激神经元。(5)评估荧光强度随时间的变化,以测量钙瞬变或突触囊泡融合。 请点击此处查看此图的放大版本。
