Моделирование бокового амиотрофического склероза (БАС) в лаборатории становится уникально сложным из-за чрезвычайно спорадического характера более 80% случаев1 в сочетании с огромным количеством генетических мутаций, которые, как известно, являются причинами заболеваний2. Несмотря на это, все случаи БАС объединяет та особенность, что до прямой нейрональной дегенерации существует дисфункциональная связь между пресинаптическими двигательными нейронами и постсинаптическими мышечными клетками3,4. Клинически, поскольку пациенты теряют связность оставшихся верхних и нижних двигательных нейронов, у них проявляются особенности гипер- и гиповозбудимости нейронов на протяжении всего заболевания5,6,7,8,9, отражающие сложные молекулярные изменения этих синапсов, которые мы, как исследователи БАС, стремимся понять.
Многочисленные трансгенные модели показали, что ухудшение и дезорганизация нервно-мышечного соединения происходит с экспрессией БАС-причинных генетических мутаций, включая SOD110, FUS11,12, C9orf7213,14,15,16 и TDP4317,18,19 посредством морфологических оценок, включая оценку синаптических бутонов, плотности позвоночника и пред/постсинаптической организации. Механически, начиная с знаковых работ Коула, Ходжкина и Хаксли в 1930-х годах, также стало возможным оценивать синаптические реакции с помощью электрофизиологических методов либо в культуре клеток in vitro, либо в препаратах среза ткани20. Благодаря этим стратегиям многие модели БАС продемонстрировали дефицит синаптической передачи. Например, мутантный вариант TDP43 вызывает повышенную частоту срабатывания и снижает порог потенциального действия в NSC-34 (спинной мозг x гибридная клеточная линия 34 нейробластомы) моторно-нейроноподобных клетках21. Этот же вариант также вызывает дисфункциональную синаптическую передачу в нервно-мышечном соединении (NMJ) до начала поведенческого двигательного дефицита у мышиной модели22. Ранее было показано, что мутантная экспрессия FUS приводит к снижению синаптической передачи в NMJ в модели дрозофилы FUS-ALS до дефектов опорно-двигательного аппарата11. Недавний отчет с использованием индуцированных плюрипотентных стволовых клеток, полученных из носителей расширения C9orf72, показал уменьшение легко высвобождаемого пула синаптических везикул23. В целом, эти и другие исследования подчеркивают важность построения более полного понимания механизмов, лежащих в основе синаптической сигнализации в моделях БАС, связанных с заболеванием. Это будет иметь решающее значение для понимания патобиологии БАС и разработки потенциальных терапевтических целей для пациентов.
Методы зажимных ячеек тока и напряжения были неоценимы при определении свойств мембраны, таких как проводимость, мембранный потенциал покоя и квантовое содержание отдельных синапсов20,24. Тем не менее, одним из существенных ограничений электрофизиологии является то, что она технически сложна и дает представление только от одного нейрона за раз. Конфокальная микроскопия живых клеток в сочетании со специфическими флуоресцентными зондами дает возможность исследовать синаптическую передачу нейронов пространственно-временным способом25,26,27. Хотя этот флуоресцентный подход не является прямой мерой возбудимости нейронов, он может обеспечить относительное измерение двух молекулярных корреляций синаптической функции: высвобождения синаптических пузырьков и переходных процессов кальция на синаптических терминалях.
Когда потенциал действия достигает пресинаптической терминальной области нейронов, запускаются переходные процессы кальция, облегчающие переход от электрического сигнала к процессу высвобождения нейромедиатора28. Кальциевые каналы с напряжением, локализованные в этих областях, жестко регулируют передачу сигналов кальция для модуляции кинетики высвобождения нейротрансмиттера29. Первые зарегистрированные флуоресцентные записи переходных процессов кальция были выполнены с использованием либо двухволнового индикатора Fura-2 AM, либо одноволнового красителя Fluo-3 AM30,31,32. Хотя эти красители предлагали большое новое понимание в то время, они страдают от нескольких ограничений, таких как неспецифическая компартментализация внутри клеток, активная или пассивная потеря красителя из меченых клеток, фотоотбеливание и токсичность при изображении в течение длительных периодов времени33. В последнее десятилетие генетически закодированные показатели кальция стали рабочими лошадками для визуализации различных форм нейронной активности. Эти показатели объединяют модифицированный флуоресцентный белок с белком хелатора кальция, который быстро переключает интенсивность флуоресценции после связывания ионов Ca2+34. Применение этих новых показателей обширно, что позволяет значительно упростить визуализацию внутриклеточных переходных процессов кальция как in vitro, так и in vivo. Одно семейство этих генетически закодированных репортеров, известное как GCaMP, в настоящее время широко используется. Эти индикаторы содержат С-концевой кальмодулиновый домен, за которым следует зеленый флуоресцентный белок (GFP), и ограничены N-концевой аллодулин-связывающей областью35,36. Связывание кальция с кальмодулиновой областью вызывает взаимодействие с кальмодулин-связывающей областью, что приводит к конформационному изменению общей структуры белка и существенному увеличению флуоресценции фрагмента GFP35,36. За прошедшие годы это семейство репортеров претерпело несколько эволюций, чтобы обеспечить четкое считывание для определенных переходных процессов кальция с определенной кинетикой (медленной, средней и быстрой), каждая из которых имеет немного разные свойства37,38. Здесь было подчеркнуто использование репортера GcaMP6, который, как было ранее показано, обнаруживает потенциалы одиночного действия и дендритные переходные процессы кальция в нейронах как in vivo, так и in vitro37.
Переходные процессы кальция в пресинаптической области вызывают события слияния синаптических пузырьков, вызывая высвобождение нейротрансмиттера в синапс и инициирование сигнальных событий в постсинаптической клетке28,39. Синаптические везикулы быстро высвобождаются и рециркулируются, так как клетка гомеостатически поддерживает стабильную площадь поверхности клеточной мембраны и легко высвобождаемый пул везикул, способных к слиянию, связанных с мембраной40. Используемый здесь стириловый краситель имеет сродство к липидным мембранам и специфически изменяет свои эмиссионные свойства в зависимости от упорядочения окружающей липидной среды41,42. Таким образом, это идеальный инструмент для маркировки рециркуляции синаптических везикул и последующего отслеживания этих везикул, поскольку они позже высвобождаются после стимуляции нейронов41,42. Протокол, который был сгенерирован и оптимизирован, является адаптацией концепций, первоначально описанных Гаффилдом и его коллегами, что позволяет нам визуализировать меченую стириловым красителем синаптикальную пузырьковую пункту с течением времени непрерывно41.
Здесь описаны две связанные флуоресцентные методологии, надежно сообщающие о конкретных клеточных событиях, участвующих в синаптической передаче. Были определены протоколы для исследования динамики деполяризационно-опосредованного пресинаптического терминального притока кальция и экзоцитоза синаптических везикул в культивируемых нейронах. Здесь методы и репрезентативные результаты сосредоточены на использовании первичных кортикальных или моторных нейронов грызунов в качестве модельной системы in vitro, поскольку опубликованы исследования с использованием этих типов клеток43,44. Тем не менее, эти методы также применимы к дифференцированным человеческим i3 кортикоподобным нейронам45, поскольку мы также добились успеха с обоими протоколами в текущих экспериментах в нашей лаборатории. Общий протокол описан в пошаговом линейном формате, показанном на рисунке 1. Короче говоря, для изучения динамики кальция в нейритах зрелые нейроны трансфектируют плазмидной ДНК для экспрессии флуоресцентного репортера GCaMP6m под промотором цитомегаловируса (ЦМВ)37,46. Трансфектированные клетки имеют низкий уровень базальной зеленой флуоресценции, которая увеличивается в присутствии кальция. Области, представляющие интерес, задаются для мониторинга изменений флуоресценции во время наших манипуляций. Это позволяет измерять высоко пространственно и временно локализованные колебания кальция37,46. Для оценки слияния и высвобождения синаптических пузырьков зрелые нейроны нагружают стириловым красителем, включенным в синаптические везикулярные мембраны, поскольку они рециркулируются, реформируются и перезаряжаются нейротрансмиттерами в пресинаптических клетках41,42,43,47,48. Современные красители, используемые для этой цели, маркируют синаптические везикулы вдоль нейритов и используются в качестве прокси для этих областей в экспериментах с живой визуализацией, как было показано совместным окрашиванием стирилового красителя и синаптотагмина Крашевским и его коллегами49. Сюда включены репрезентативные изображения аналогичного окрашивания, которые также были выполнены (рисунок 2А). Предыдущие исследователи широко использовали такие красители, чтобы сообщить о динамике синаптических пузырьков в нервно-мышечном соединении и нейронах гиппокампа48,49,50,51,52,53,54,55,56 . Путем выбора пунктатных областей везикул, нагруженных красителем, и мониторинга снижения интенсивности флуоресценции после высвобождения везикул можно изучить функциональную синаптическую пропускную способность и временную динамику высвобождения после стимуляции43. Для обоих способов среда, содержащая высокую концентрацию хлорида калия, используется для деполяризации клеток для имитации активности нейронов. Параметры визуализации задаются для захвата интервалов менее секунды, охватывающих базовую нормализацию, за которой следует наш период захвата стимуляции. Измерения флуоресценции в каждой точке времени определяются, нормализуются на заднем плане и количественно оцениваются в течение экспериментального периода времени. С помощью этой стратегии можно обнаружить опосредованное притоком кальция флуоресценцию GCaMP6m или эффективное снижение флуоресценции высвобождения синаптических везикул экзоцитоза стириловых красителей. Ниже описываются подробные методологические настройки и параметры этих двух протоколов, а также обсуждение их преимуществ и ограничений.
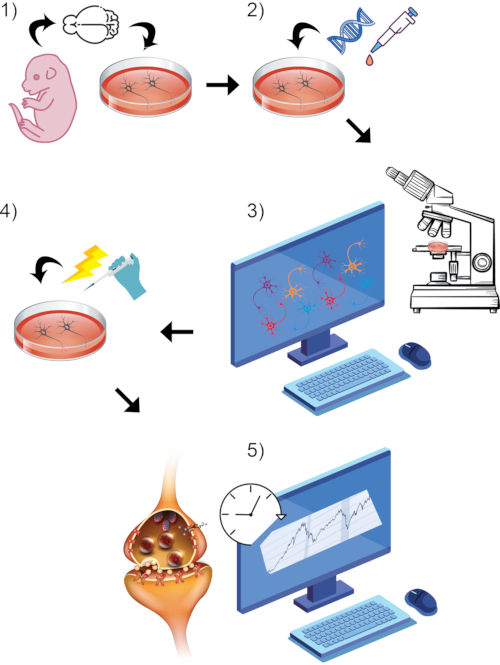
Рисунок 1: Визуальный рендеринг всего процесса общего протокола. (1) Изолировать и культивировать первичные нейроны грызунов in vitro до выбранной временной точки созревания. (2) Ввести ДНК GCaMP или стириловый краситель в качестве репортеров синаптической активности. (3) Настройка парадигмы визуализации с использованием конфокального микроскопа, оснащенного живой визуализацией, и связанного с ним программного обеспечения. Начните базовый период записи. (4) В то время как клетки все еще подвергаются захвату живого изображения, стимулируйте нейроны с помощью перфузии ванны с высоким KCl. (5) Оценка измерений интенсивности флуоресценции с течением времени для измерения переходных процессов кальция или синаптического слияния пузырьков. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
