प्रयोगशाला में मॉडलिंग एमियोट्रोफिक लेटरल स्केलेरोसिस (एएलएस) को 80% से अधिक मामलों की भारी छिटपुट प्रकृति के कारण विशिष्ट रूप से चुनौतीपूर्ण बनाया गया है1, जो रोग-प्रेरक 2 के रूप में जाने जाने वाले आनुवंशिक उत्परिवर्तन की विशाल संख्या के साथ युग्मित है। इसके बावजूद, एएलएस के सभी मामले एकीकृत विशेषता को साझा करते हैं कि एकमुश्त न्यूरोनल अध: पतन से पहले, प्रीसिनेप्टिक मोटर न्यूरॉन्स और पोस्टसिनेप्टिक मांसपेशी कोशिकाओं के बीच बेकार संचार होता है3,4। नैदानिक रूप से, जैसा कि रोगी शेष ऊपरी और निचले मोटर न्यूरॉन्स की कनेक्टिविटी खो देते हैं, वे पूरे रोग 5,6,7,8,9 में न्यूरोनल हाइपर- और हाइपोएक्साइटबिलिटी की विशेषताओं के साथ पेश करते हैं, जो इन synapses में जटिल अंतर्निहित आणविक परिवर्तनों को दर्शाते हैं, जिसे हम, एएलएस शोधकर्ताओं के रूप में, समझना चाहते हैं।
एकाधिक ट्रांसजेनिक मॉडल ने सचित्र किया है कि न्यूरोमस्कुलर जंक्शन की गिरावट और अव्यवस्था एएलएस-प्रेरक आनुवंशिक उत्परिवर्तन की अभिव्यक्ति के साथ होती है, जिसमें SOD110, FUS11,12, C9orf7213,14,15,16, और TDP4317,18,19 शामिल हैं। रूपात्मक मूल्यांकन के माध्यम से, जिसमें सिनैप्टिक बाउटन, रीढ़ की हड्डी के घनत्व और पूर्व / पोस्टसिनेप्टिक संगठन का मूल्यांकन शामिल है। यंत्रवत रूप से, 1 9 30 के दशक में कोल, हॉजकिन और हक्सले के ऐतिहासिक कागजात के बाद से, इन विट्रो सेल संस्कृति या ऊतक स्लाइस तैयारी में इलेक्ट्रोफिजियोलॉजिकल तकनीकों के माध्यम से सिनैप्टिक प्रतिक्रियाओं का मूल्यांकन करना भी संभव हो गया है। इन रणनीतियों के माध्यम से, एएलएस के कई मॉडलों ने सिनैप्टिक ट्रांसमिशन घाटे का प्रदर्शन किया है। उदाहरण के लिए, TDP43 का एक उत्परिवर्ती संस्करण बढ़ी हुई फायरिंग आवृत्ति का कारण बनता है और एनएससी -34 (रीढ़ की हड्डी एक्स न्यूरोब्लास्टोमा हाइब्रिड सेल लाइन 34) मोटर-न्यूरॉन जैसी कोशिकाओं 21 में एक्शन पोटेंशियल थ्रेशोल्ड को कम करता है। यह एक ही संस्करण भी एक माउस model22 में व्यवहार मोटर घाटे की शुरुआत से पहले neuromuscular जंक्शन (NMJ) पर बेकार synaptic संचरण का कारण बनता है। यह पहले दिखाया गया था कि उत्परिवर्ती एफयूएस अभिव्यक्ति के परिणामस्वरूप लोकोमोटर दोष 11 से पहले एफयूएस-एएलएस के ड्रोसोफिला मॉडल में एनएमजे पर कम सिनैप्टिक संचरण होता है। C9orf72-विस्तार वाहक से व्युत्पन्न प्रेरित प्लुरिपोटेंट स्टेम कोशिकाओं का उपयोग करके हाल ही में एक रिपोर्ट ने सिनैप्टिक पुटिकाओं 23 के आसानी से पुनरावृत्ति योग्य पूल में कमी का खुलासा किया। कुल मिलाकर, ये अध्ययन और अन्य एएलएस के रोग-प्रासंगिक मॉडल में सिनैप्टिक सिग्नलिंग के अंतर्निहित तंत्र की अधिक व्यापक समझ के निर्माण के महत्व को उजागर करते हैं। यह एएलएस के पैथोबायोलॉजी को समझने और रोगियों के लिए संभावित चिकित्सीय लक्ष्यों को विकसित करने में महत्वपूर्ण होगा।
वर्तमान और वोल्टेज क्लैंपिंग कोशिकाओं के तरीके झिल्ली गुणों को निर्धारित करने में अमूल्य रहे हैं जैसे कि चालकता, आराम झिल्ली क्षमता, और व्यक्तिगत synapses20,24 की क्वांटल सामग्री। हालांकि, इलेक्ट्रोफिजियोलॉजी की महत्वपूर्ण सीमाओं में से एक यह है कि यह तकनीकी रूप से चुनौतीपूर्ण है और केवल एक समय में एक ही न्यूरॉन से अंतर्दृष्टि प्रदान करता है। लाइव-सेल कॉन्फोकल माइक्रोस्कोपी, विशिष्ट फ्लोरोसेंट जांच के साथ युग्मित, न्यूरॉन्स के सिनैप्टिक ट्रांसमिशन की जांच करने का अवसर प्रदान करता है एक स्पैटिओटेम्पोरल तरीके से 25,26,27। हालांकि न्यूरोनल उत्तेजना का एक प्रत्यक्ष उपाय नहीं है, यह प्रतिदीप्ति दृष्टिकोण सिनैप्टिक फ़ंक्शन के दो आणविक सहसंबंधों का एक सापेक्ष माप प्रदान कर सकता है: सिनैप्टिक पुटिका रिलीज और सिनैप्टिक टर्मिनलों पर कैल्शियम क्षणिक।
जब एक कार्रवाई क्षमता न्यूरॉन्स के प्रीसिनेप्टिक टर्मिनल क्षेत्र तक पहुंचती है, तो कैल्शियम क्षणिकों को ट्रिगर किया जाता है, जिससे न्यूरोट्रांसमीटर रिलीज 28 की प्रक्रिया में विद्युत संकेत से संक्रमण की सुविधा मिलती है। वोल्टेज-गेटेड कैल्शियम चैनल इन क्षेत्रों के लिए स्थानीयकृत कसकर कैल्शियम सिग्नलिंग को विनियमित करने के लिए न्यूरोट्रांसमीटर रिलीज 29 के कैनेटीक्स को संशोधित करने के लिए। कैल्शियम transients की पहली रिपोर्ट प्रतिदीप्ति आधारित रिकॉर्डिंग या तो दोहरी तरंग दैर्ध्य संकेतक Fura-2 AM या एकल तरंग दैर्ध्य डाई Fluo-3 AM30,31,32 का उपयोग कर प्रदर्शन किया गया था। जबकि इन रंजकों ने उस समय महान नई अंतर्दृष्टि की पेशकश की थी, वे कई सीमाओं से पीड़ित हैं जैसे कि कोशिकाओं के भीतर गैर-विशिष्ट कंपार्टमेंटलाइजेशन, लेबल कोशिकाओं से सक्रिय या निष्क्रिय डाई हानि, फोटोब्लीचिंग और विषाक्तता यदि समय 33 की विस्तारित अवधि में चित्रित किया जाता है। पिछले दशक में, आनुवंशिक रूप से एन्कोडेड कैल्शियम संकेतक न्यूरोनल गतिविधि के विभिन्न रूपों की इमेजिंग के लिए वर्कहॉर्स बन गए हैं। ये संकेतक कैल्शियम चेलेटर प्रोटीन के साथ एक संशोधित फ्लोरोसेंट प्रोटीन को जोड़ते हैं जो Ca2 + आयन34 के बंधन के बाद तेजी से प्रतिदीप्ति तीव्रता को स्विच करता है। इन नए संकेतकों का आवेदन विशाल है, जो इन विट्रो और विवो सेटिंग्स दोनों में इंट्रासेल्युलर कैल्शियम क्षणिकों के बहुत आसान विज़ुअलाइज़ेशन की अनुमति देता है। इन आनुवंशिक रूप से एन्कोडेड पत्रकारों का एक परिवार, जिसे जीसीएएमपी के रूप में जाना जाता है, अब व्यापक रूप से उपयोग किया जाता है। इन संकेतकों में एक सी-टर्मिनल कैलमोडुलिन डोमेन होता है, जिसके बाद हरे रंग के फ्लोरोसेंट प्रोटीन (जीएफपी) होते हैं, और एक एन-टर्मिनल कैलमोडुलिन-बाइंडिंग क्षेत्र 35,36 द्वारा कैप किए जाते हैं। कैलमोडुलिन डोमेन के लिए कैल्शियम-बाइंडिंग कैलमोडुलिन-बाइंडिंग क्षेत्र के साथ एक बातचीत को ट्रिगर करता है, जिसके परिणामस्वरूप समग्र प्रोटीन संरचना में एक संरचनात्मक परिवर्तन होता है और जीएफपी समूह 35,36 के प्रतिदीप्ति में पर्याप्त वृद्धि होती है। इन वर्षों में, पत्रकारों के इस परिवार ने विशिष्ट कैनेटीक्स (धीमी गति से, मध्यम और तेज) के साथ विशेष कैल्शियम क्षणिकों के लिए अलग-अलग रीडआउट को सक्षम करने के लिए कई विकास किए हैं, जिनमें से प्रत्येक में थोड़ा अलग गुण हैं37,38। यहां, रिपोर्टर GcaMP6 के उपयोग पर प्रकाश डाला गया है, जिसे पहले विवो और इन विट्रो 37 दोनों में न्यूरॉन्स में एकल एक्शन पोटेंशियल और डेंड्राइटिक कैल्शियम क्षणिकों का पता लगाने के लिए दिखाया गया है।
प्रीसिनेप्टिक क्षेत्र में कैल्शियम क्षणिक सिनैप्टिक पुटिका संलयन घटनाओं को ट्रिगर करते हैं, जिससे न्यूरोट्रांसमीटर रिलीज होता है सिनैप्स में और पोस्टसिनेप्टिक सेल 28,39 में सिग्नलिंग घटनाओं की दीक्षा होती है। सिनैप्टिक पुटिकाओं को तेजी से जारी और पुनर्नवीनीकरण दोनों किया जाता है, क्योंकि सेल होमोस्टैटिक रूप से एक स्थिर कोशिका झिल्ली सतह क्षेत्र को बनाए रखता है और संलयन सक्षम झिल्ली-बाध्य पुटिकाओं के आसानी से पुन: प्रयोज्य पूल 40 को बनाए रखता है। यहां उपयोग किए जाने वाले स्टायरिल डाई में लिपिड झिल्ली के प्रति एक आत्मीयता है और विशेष रूप से आसपास के लिपिड वातावरण 41,42 के आदेश के आधार पर इसके उत्सर्जन गुणों को बदलता है। इस प्रकार, यह रीसाइक्लिंग सिनैप्टिक पुटिकाओं को लेबल करने और इन पुटिकाओं के बाद के ट्रैकिंग के लिए एक आदर्श उपकरण है क्योंकि वे बाद में न्यूरोनल उत्तेजना 41,42 के बाद जारी किए जाते हैं। प्रोटोकॉल जो उत्पन्न और अनुकूलित किया गया है, वह गफफील्ड और सहकर्मियों द्वारा शुरू में वर्णित अवधारणाओं का एक अनुकूलन है, जो हमें समय के साथ स्टाइरिल डाई-लेबल वाले सिनैप्टिक पुटिका पंक्टा की कल्पना करने की अनुमति देता है।
यहां, दो संबंधित प्रतिदीप्ति-आधारित तरीकों का वर्णन किया गया है, मज़बूती से सिनैप्टिक ट्रांसमिशन में शामिल विशिष्ट सेलुलर घटनाओं की रिपोर्टिंग। सुसंस्कृत न्यूरॉन्स में विध्रुवीकरण-मध्यस्थता वाले प्रीसिनेप्टिक टर्मिनल कैल्शियम प्रवाह और सिनैप्टिक पुटिका एक्सोसाइटोसिस की गतिशीलता की जांच करने के लिए प्रोटोकॉल को परिभाषित किया गया है। यहां, विधियों और प्रतिनिधि परिणामों को इन विट्रो मॉडल सिस्टम के रूप में प्राथमिक कृंतक कॉर्टिकल या मोटर न्यूरॉन्स का उपयोग करने पर ध्यान केंद्रित किया जाता है, क्योंकि इन सेल प्रकारों का उपयोग करके प्रकाशित अध्ययन हैं43,44। हालांकि, ये विधियां विभेदित मानव i3 कॉर्टिकल-जैसे न्यूरॉन्स 45 पर भी लागू होती हैं, क्योंकि हमें अपनी प्रयोगशाला में वर्तमान में चल रहे प्रयोगों में दोनों प्रोटोकॉल के साथ भी सफलता मिली है। सामान्य प्रोटोकॉल को चरणबद्ध रैखिक प्रारूप में रेखांकित किया गया है, जो चित्र 1 में दिखाया गया है। संक्षेप में, न्यूराइट्स में कैल्शियम गतिशीलता का अध्ययन करने के लिए, परिपक्व न्यूरॉन्स को एक साइटोमेगालोवायरस (सीएमवी) प्रमोटर 37,46 के तहत फ्लोरोसेंट रिपोर्टर जीसीएएमपी 6 एम को व्यक्त करने के लिए प्लास्मिड डीएनए के साथ संक्रमित किया जाता है। ट्रांसफेक्टेड कोशिकाओं में बेसल ग्रीन प्रतिदीप्ति का निम्न स्तर होता है, जो कैल्शियम की उपस्थिति में बढ़ता है। ब्याज के क्षेत्रों को हमारे हेरफेर के दौरान प्रतिदीप्ति परिवर्तनों की निगरानी करने के लिए निर्दिष्ट किया जाता है। यह कैल्शियम में अत्यधिक स्थानिक और अस्थायी रूप से स्थानीयकृत उतार-चढ़ाव को मापा जा सकता है37,46। सिनैप्टिक पुटिका संलयन और रिलीज का मूल्यांकन करने के लिए, परिपक्व न्यूरॉन्स को सिनैप्टिक पुटिका झिल्ली में शामिल स्टाइरिल डाई के साथ लोड किया जाता है क्योंकि वे प्रीसिनेप्टिक कोशिकाओं में न्यूरोट्रांसमीटर के साथ पुनर्नवीनीकरण, सुधार और पुनः लोड किए जाते हैं41,42,43,47,48। इस उद्देश्य के लिए उपयोग किए जाने वाले वर्तमान रंजक न्यूराइट्स के साथ सिनैप्टिक पुटिकाओं को लेबल करते हैं और लाइव-इमेजिंग प्रयोगों में इन क्षेत्रों के लिए एक प्रॉक्सी के रूप में उपयोग किए जाते हैं, जैसा कि क्राज़ेवस्की और सहकर्मियों द्वारा स्टाइरिल डाई और सिनैप्टोटाग्मिन के सह-धुंधला द्वारा दिखाया गया था। यहां शामिल हैं इसी तरह के दाग की प्रतिनिधि छवियां जो भी की गई हैं (चित्रा 2 ए)। पिछले जांचकर्ताओं ने न्यूरोमस्कुलर जंक्शन और हिप्पोकैम्पल न्यूरॉन्स 48,49,50,51,52,53,54,55,56 पर सिनैप्टिक पुटिका गतिशीलता की रिपोर्ट करने के लिए बड़े पैमाने पर इस तरह के रंगों का उपयोग किया है . डाई-लोडेड पुटिकाओं के पंक्टेट क्षेत्रों का चयन करके और पुटिका रिलीज के बाद प्रतिदीप्ति तीव्रता में कमी की निगरानी करके, कार्यात्मक सिनैप्टिक ट्रांसमिशन क्षमता और रिहाई की अस्थायी गतिशीलता का अध्ययन उत्तेजना 43 के बाद किया जा सकता है। दोनों विधियों के लिए, पोटेशियम क्लोराइड की उच्च सांद्रता वाले एक माध्यम को न्यूरोनल गतिविधि की नकल करने के लिए कोशिकाओं को विध्रुवीकृत करने के लिए नियोजित किया जाता है। इमेजिंग पैरामीटर को हमारे उत्तेजना कैप्चर अवधि के बाद बेसलाइन सामान्यीकरण में फैले उप-दूसरे अंतराल को कैप्चर करने के लिए निर्दिष्ट किया जाता है। प्रत्येक समय बिंदु पर प्रतिदीप्ति माप निर्धारित किए जाते हैं, पृष्ठभूमि के लिए सामान्यीकृत होते हैं, और प्रयोगात्मक समय अवधि में परिमाणित होते हैं। कैल्शियम-प्रवाह मध्यस्थता GCaMP6m प्रतिदीप्ति वृद्धि या प्रभावी synaptic पुटिका exocytosis styryl डाई रिलीज प्रतिदीप्ति कमी इस रणनीति के माध्यम से पता लगाया जा सकता है। इन दो प्रोटोकॉल के लिए विस्तृत पद्धतिगत सेटअप और पैरामीटर और उनके लाभों और सीमाओं पर चर्चा नीचे वर्णित हैं।
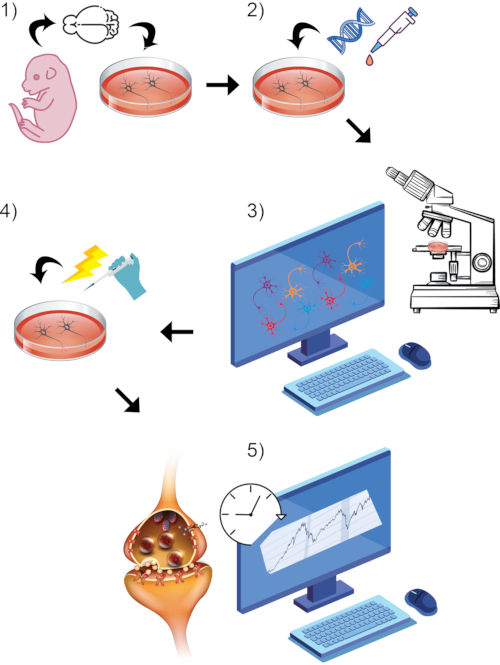
चित्रा 1: समग्र सामान्य प्रोटोकॉल प्रक्रिया का दृश्य रेंडरिंग। (1) पृथक और संस्कृति प्राथमिक कृंतक न्यूरॉन्स विट्रो में चुना परिपक्वता timepoint करने के लिए. (2) GCaMP डीएनए या स्टाइरिल डाई को सिनैप्टिक गतिविधि के पत्रकारों के रूप में पेश करें। (3) लाइव इमेजिंग सुसज्जित confocal माइक्रोस्कोप और संबंधित सॉफ्टवेयर का उपयोग कर सेटअप इमेजिंग प्रतिमान. बेसलाइन रिकॉर्डिंग अवधि प्रारंभ करें. (4) जबकि कोशिकाएं अभी भी लाइव-इमेज कैप्चर से गुजर रही हैं, उच्च केसीएल स्नान परफ्यूजन के माध्यम से न्यूरॉन्स को उत्तेजित करती हैं। (5) कैल्शियम क्षणिक या सिनैप्टिक पुटिका संलयन को मापने के लिए समय के साथ प्रतिदीप्ति तीव्रता माप का आकलन करें। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहाँ क्लिक करें.
