This section shows the workflow of an ex vivo optogenetic approach and representative results from different experimental strategies to investigate the physiological properties of sensory and modulatory long-range projections to BLA and mpITC neurons as well as properties of local connectivity between mpITC and BLA.
After stereotactic injection of the selected viral vector at the desired coordinates into the mouse brain (Figure 1A-C, viral expression time 2 – 6 weeks, depending on experiment), acute brain slices of the injection sites and projection areas in the amygdala are prepared at the appropriate angle for patch-clamp experiments and from the injected brain region for post-hoc analysis of injection sites (Figure 1D). Before starting recordings and the optogenetic stimulation, fluorescently labeled cells or axonal projections in the target area (amygdala) should be checked on the upright microscope for patch-clamp recordings by using the attached fluorescent lamp (Figure 1E). Upon obtaining a patch-clamp recording from a putative target cell in the projection region of the slice, light stimulation is initiated while time point, interval, and pulse length are controlled via the patch software that triggers the light emitting diode (LED). Depending on the experimental strategy (Figure 2A-C, right) the aperture in the fluorescent light path is either fully opened or restricted (Figure 2D). While keeping the pulse length constant and as short as possible (ideally ≤1 msec), LED output intensity is slowly increased to assess which intensity is required to achieve the desired amplitude of the synaptic response in a particular experiment (Figure 2E). After recording, amygdala slices are post-fixed. Upon resectioning and optional staining, injection and recording sites are imaged on a confocal microscope to verify injection location and to exclude data from misplaced injections. Images of injection sites and the associated axonal projections in the amygdala for the three experimental strategies (mPFC inputs to BLA, thalamic inputs to mpITCs, and local mpITC activation) are shown in Figure 2A-C.
Figure 3 shows representative recording results obtained from BLA principal neurons, local BLA interneurons and mpITC neurons illustrating the properties of light-evoked responses for the different experimental strategies used (Figure 3A, E, I). To target GABAergic neurons (local interneurons and mpITCs) for recording, GAD67-GFP reporter mice were used 24. For mPFC and sensory thalamic projections, several aspects of synaptic transmission can be studied. The temporal precision of activation and the kinetic properties of the ChRs used in this study enable reliable stimulation with paired pulses at an inter-stimulus-interval of 50 msec. This allows for analysis of paired-pulse ratios (PPR) of postsynaptic currents (PSCs), which can either be facilitating or depressing depending on projection and target cell type, and serve as indicators of presynaptic release probability (Figure 3B, F, H). Additionally, analysis of synaptic latencies allows for dissection of different postsynaptic response components.
In this example, longer latencies of the inhibitory PSC (IPSC) versus excitatory PSC (EPSC) component indicate a disynaptic and monosynaptic input, respectively (Figure 3C). In other instances, a more thorough analysis of additional EPSC properties such as response jitter or the coefficient of variance of response size may be needed to draw conclusions on its mono- or disynaptic nature 17,23. Furthermore, application of pharmacological blockers for specific receptors can identify the nature of PSCs (i.e., glutamatergic EPSC and GABAergic IPSC, Figure 3C-D). As expected, the early component of the mPFC input was glutamatergic, whereas the late component was GABAergic. The complete block of both EPSC and IPSC with the glutamate receptor blocker CNQX further supports that the IPSC is disynaptic. To investigate modulation of synaptic transmission at optogenetically activated fibers, the effects of agonists and antagonists for metabotropic receptors (here GABAB receptors) on amplitude and PPR can be assessed to evaluate influences on pre- and postsynaptic sites. In this example, the concomitant decrease of amplitude with an increase in PPR is indicative of a presynaptic modulation of light-activated fibers (3G, H). Finally, local interactions of cells in the amygdala can be assessed, for example, when mice that express Cre under control of the Tac2 promoter 25 are injected with a double-floxed ChR expressing viral vector (Figure 1C). Because Tac2 and thus, ChR is expressed in mpITC and central amygdala (Figure 2C), light activation was restricted to the mpITC by closing the fluorescent light path aperture (Figure 2D). Under these conditions, light-evoked action potentials can be elicited in infected mpITCs. Short latency inhibitory synaptic responses in the BLA indicate the presence of a functional inhibitory connection between mpITC and BLA principal neurons.
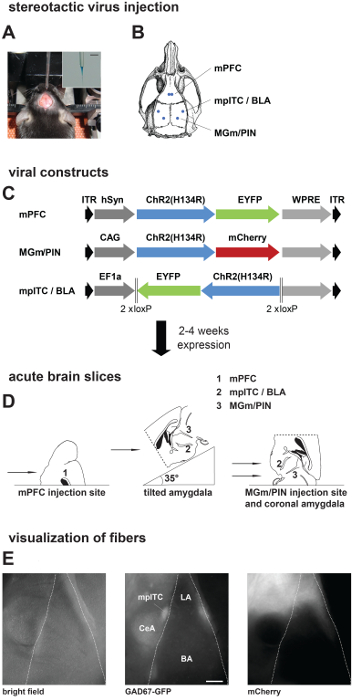
Figure 1. Stereotactic Injections, Preparation of Acute Brain Slices, and Visualization of Presynaptic Fibers. (A, B) Stereotactic virus injection. A) Picture of anesthetized mouse placed in a stereotactic frame with skull exposed and the injection pipette. Inset: Zoom in picture of injection pipette filled with virus solution mixed with fast green. Scale bar: 3 mm. (B) Schematic of a mouse skull with marked positions of drill holes for different injection areas. (C) Scheme showing different viral constructs used in this study. Dark grey: promoter sequence; blue: Channelrhodopsin2 (ChR2 (H134R)); green/red: fluorescent protein. Expression time was 2 weeks for local amygdala projections and 4 – 6 weeks for projections from mPFC and MGm/PIN. (D) Preparation of acute brain slices: Scheme showing placement of mouse brain on slicer stage for obtaining slices from different injection and projection areas. (E) Visualization of fibers in acute brain slices from a GAD67-GFP mouse injected with ChR2-mCherry virus in MGm/PIN. Pictures are taken on the upright patch microscope with different filter sets: GAD67-GFP expression, middle; MGm/PIN fibers labeled with mCherry, right. Scale bar: 200 µm. mPFC, medial prefrontal cortex; mpITC, medial paracapsular intercalated cells; BLA, basolateral amygdala; MGm, medial geniculate nucleus, medial part; PIN, posterior intralaminar thalamic nucleus; LA, lateral amygdala; BA, basal amygdala; CeA, central nucleus of the amygdala. Please click here to view a larger version of this figure.
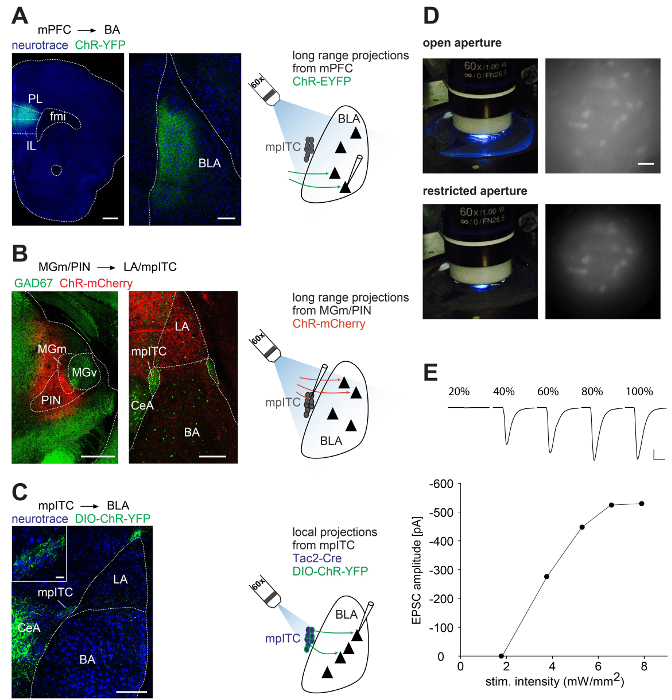
Figure 2. Injection Sites, Projection Areas, and Optogenetic Stimulation of Axonal Projections. (A-C) Confocal images of representative injection sites and projection areas in the BLA, and schemes of the experimental strategies. (A) Left: mPFC injection site in a C57Bl/6 mouse stained with Neurotrace. Scale bar: 500 µm. Middle: corresponding projections in the BLA in a tilted amygdala slice. Scale bar: 200 µm. Right: Schematic of whole field illumination of mPFC projections and recording of neuron in the BLA. (B) Left: MGm/PIN injection site in a GAD67-GFP mouse. Scale bar: 500 µm. Middle: Corresponding projections in mpITC and LA of a coronal amygdala slice. Scale bar: 200 µm. Right: Schematic of whole field illumination of MGm/PIN projections and recording of an mpITC neuron. (C) Left: Local expression of ChR2-YFP in a Neurotrace stained amygdala slice of a Tac2-Cre mouse. Scale bar: 200 µm. Inset: Zoom in of mpITCs expressing ChR2-YFP. Scale bar: 20 µm. Right: Schematic of restricted illumination of the mpITC cluster and recording of a BLA neuron. (D) Pictures of recording chamber during light delivery and image of neurons in acute brain slices (mpITC cluster) in a GAD67-GFP mouse with open and restricted aperture. Scale bar: 30 µm. (E) Excitatory postsynaptic currents (EPSCs) evoked by different LED intensities (top) and plot of light power versus evoked EPSC amplitude (bottom) from a representative mpITC neuron upon optogenetic activation of fibers from MGm/PIN. Scale bar: 100 pA/10 msec.
Note: Figure 2A is modified from reference #16 and Figure 2C from reference #23. Please click here to view a larger version of this figure.
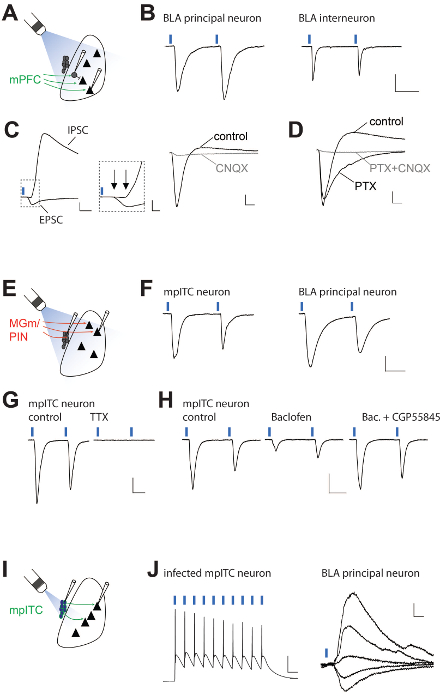
Figure 3. Exemplary Results from Optogenetic Activation of Long Range and Local Projections. (A) Scheme of experimental strategy for (B-D). (B) Representative EPSCs recorded from a BLA principal neuron and BLA interneuron upon optogenetic paired-pulse stimulation (50 msec inter-stimulus-interval) of fibers from mPFC eliciting paired pulse facilitation and paired pulse depression, respectively. Scale bar: 100 pA/25 msec. (C) Optogenetic activation of fibers from mPFC elicits feed-forward inhibition. Left: Representative EPSC (at -70 mV) and inhibitory postsynaptic current (IPSC, at 0 mV) in a BLA principal neuron. The IPSC has a longer synaptic latency compared to the EPSC. Scale bars: 200 pA/10 msec and 200 pA/2 msec. Right: Light evoked biphasic EPSC/IPSC sequence (at -50 mV) is blocked by the AMPA/Kainate antagonist CNQX (10 µm), further supporting the disynaptic nature of the IPSC. Scale bar: 50 pA/5 msec. (D) Effects of subsequent block of EPSC/IPSC sequence (at -50 mV) by the Cl– channel blocker Picrotoxin (PTX, 100 µM) and PTX + CNQX. The IPSC is blocked by PTX and the remaining EPSC by CNQX. Scale bars: 50 pA/5 msec. (E) Scheme of experimental strategy for (F-H). F) Representative EPSCs recorded from an mpITC and a BLA principal neuron upon optogenetic paired-pulse stimulation of fibers from MGm/PIN. Scale bar: 50 pA/20 msec. (G) EPSCs in another mpITC neuron are blocked by the sodium channel blocker Tetrodotoxin (TTX, 0.5 µM), indicating that they are dependent on sodium channel activity. Scale bar: 50 pA/20 msec. (H) Thalamic inputs to mpITC neurons are modulated by presynaptic GABAB receptors. EPSCs under control condition, reduction of EPSC amplitude and concomitant increase in the paired pulse ratio during application of the GABAB agonist Baclofen (2 µM), and recovery of both amplitude and initial paired pulse ration upon co-application of the GABAB antagonist CGP55845 (10 µM). These changes are indicative of a presynaptic modulation by GABAB receptors. Scale bar: 50 pA/20 msec. (I) Scheme of experimental strategy for (J). (J) Light-evoked action potentials recorded from a ChR2-YFP expressing mpITC neuron. Light-evoked IPSCs recorded from a BLA principal neuron at different holding potentials (-90, -70, -50, -20, and 0 mV) reverse around the calculated equilibrium potential for Cl–. Scale bars: 20 mV/100 msec and 10 pA/10 msec. Note: Figure 3B-D is modified from Reference #16, and Figure 3H and J from reference #23. Please click here to view a larger version of this figure.
