To demonstrate the potential of the eyFLP-MARCM technique to generate GFP-marked patches of defined genotypes in Drosophila EAD, three types of clones were induced: (1) control expressing GFP only, (2) malignant tumors expressing an oncogenic form of the small G-protein Ras (RasV12) in a background of homozygous loss of a tumor suppressor gene scribble (scrib1), and (3) overgrowing but non-invasive rasV12scrib1jnkDN clones, where the Jun-N-terminal kinase (JNK) was inactivated by expression of its dominant-negative form (Figure 4, Table 1 for genotypes). It has been shown that the invasiveness of rasV12scrib1 clonal tumors requires aberrant activation of JNK signaling and its downstream transcription factors 10,12,23,24. Moreover, rasV12scrib1 tumors promote a strong inflammatory response in the Drosophila larva, resulting in infiltration of immune cells (called hemocytes) into the EAD 14,25.
To monitor the levels and spatial distribution of JNK activity within the mosaic tissue and among tumors of distinct genotypes, the established transgenic, JNK-responsive TRE-DsRed reporter was employed 26. Confocal microscopy of the dissected, fixed EAD and EAD/brain complexes revealed marked upregulation of the TRE-DsRed reporter activity in the tissues bearing malignant rasV12scrib1 tumors (Figure 4B). The DsRed signal labeled rasV12scrib1 clones in EAD (Figure 4B) as well as tumor cells spreading over the brain lobes and invading the VNC (Figure 4E, 4F). In contrast, the TRE-DsRed reporter activity remained restricted to a string of cells extending from the antennal to the eye part of the control discs (Figure 4A'). Blocking JNK signaling resulted in a dramatic decrease of TRE-DsRed signal in clonal rasV12scrib1jnkDN tumors (Figure 4C, 4D). These data thus provide functional evidence for a requirement of JNK signaling to activate the TRE-dependent transcriptional response in malignant rasV12scrib1 tumors. A lineage-specific hmlΔ-DsRed reporter, devised for tracing Drosophila hemocytes 27,28 showed an accumulation of immune cells in tissues bearing malignant rasV12scrib1 tumors (Figure 5B). In contrast, only individual hemocytes or a few localized hemocyte patches were detected in control and rasV12scrib1jnkDN mosaic EAD and brain tissues (Figure 5A, 5C). Immunostaining with a pan-hemocyte anti-Hemese antibody (H2) 29 confirmed the immune cell identity of hmlΔ-DsRed positive cells (Figure 5D, 5E). Consistent with published reports 12,23,24, unbiased quantification of tumor invasiveness corroborated a central role of JNK signaling in promoting tumor invasion into the adjacent brain lobes and VNC (Figure 3C). Finally, total RNA was isolated from the mosaic EAD. Samples were subjected either to qRT-PCR or analyzed on an automated electrophoresis system for quality and quantity. Figure 6A shows significant JNK-dependent upregulation of matrix metalloproteinase 1 (mmp1) transcript in EAD bearing malignant rasV12scrib1 clonal tumors, confirming the previously published findings 12,24,30,31. Assessment of total RNA using an automated electrophoresis system showed that three independent EAD samples had a 28S/18S ratio above 1.8 (Figure 6B). However, EAD 2 RNA was partially degraded, reflected by a smear on the gel (Figure 6C). In contrast, EAD 3 sample had a low concentration. Multiple bands of higher molecular weight on gel image (Figure 6B) and small peaks on the electropherogram (Figure 6C), also suggested DNA contamination. Thus, only EAD 1 sample would be suitable for a preparation of mRNA-seq library.
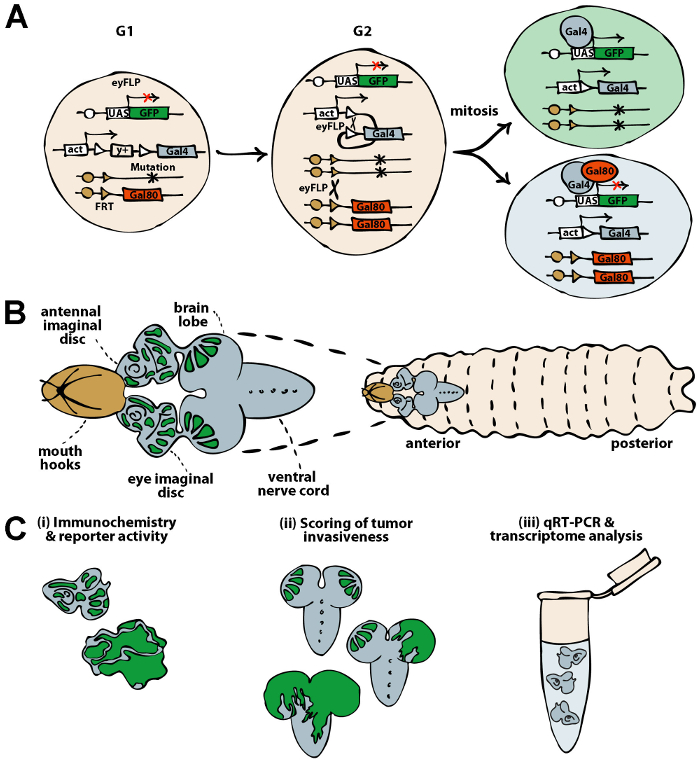
Figure 1: Schematics of the MARCM system for generation of genetic mosaics in the Drosophila EAD and downstream applications. (A) The MARCM system enables generation of clonal patches with defined genetic lesions in an otherwise wild type (heterozygous) context. The expression of FLP under the control of a tissue specific promoter (e.g., eyeless) catalyzes recombination between the two FRT elements flanking the STOP cassette marked with a yellow (y) gene (act>y+>Gal4). Upon removal of the STOP cassette, the ubiquitously expressed Gal4 transcriptional activator (act-Gal4) can drive expression of any UAS-based transgene such as UAS-GFP (also UAS-rasV12). In a parental cell, however, the Gal4 activity is blocked by a Gal80 repressor whose expression is controlled by a tubulin promoter (tub-Gal80). In the G2 phase of the cell cycle, FLP mediates the exchange of the non-sister chromatids between the homologous chromosomes distal to the FRT sites. The segregation of the recombinant chromosomes during mitosis will give rise to two daughter cells, one being homozygous mutant for a particular genomic locus (e.g., scrib1) while the other will carry two wild type alleles. Loss of Gal80 in the homozygous mutant cells will permit expression of GFP. In contrast, a wild type sister stays GFP-negative. (B) A schematic drawing of a third instar larva depicts the EAD pair connected anteriorly to the mouth hooks and posteriorly to the brain via optic stalks. The eyFLP-MARCM system induces GFP-marked clones within the EAD and the neuroepithelium of the brain lobes. (C) Dissected EAD and brains can be subjected to diverse downstream applications including immunochemistry and detection of transgenic reporter activity (i), quantification of tumor invasiveness (ii) and transcriptome profiling using a candidate (qRT-PCR) or an unbiased approach (mRNA-seq) (iii). Please click here to view a larger version of this figure.
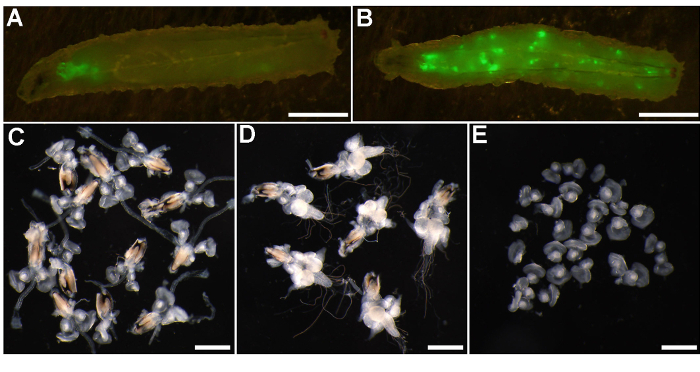
Figure 2: Sorting of third instar larvae and representative examples of EAD and EAD/brain complexes used for different downstream applications. (A-B) Fluorescent micrographs of the third instar larvae bearing control GFP-labeled clones induced with the eyFLP-MARCM 82B Green tester. (A) Representative control larva with clones restricted to the EAD and brain lobes. (B) An example of a "leopard larva" carrying GFP-positive clones in various tissues throughout the body. (C) Brightfield images of dissected EAD and (D) EAD/brain complexes attached to the mouth hooks, and (E) EAD cleaned from all extraneous tissue including mouth hooks. Scale bars = 2 mm (A-B) and 500 µm (C-E). Please click here to view a larger version of this figure.
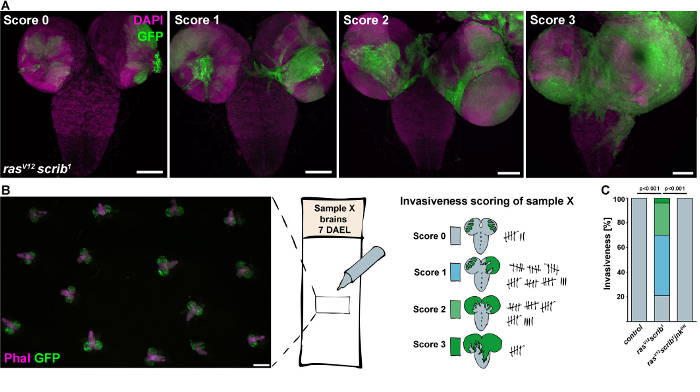
Figure 3: Quantification of tumor invasiveness. (A) Fluorescent confocal images of brains dissected from rasV12 scrib1 larvae (7 days AEL) represent the real examples of the four different grades of tumor invasiveness ranging from non-invasive (Score 0), one brain lobe invaded (Score 1), both brain lobes invaded (Score 2) to a strong tumor invasion with clonal tissue covering both brain lobes and entering the VNC (Score 3). Images are projections of multiple confocal sections and examples for Score 2 and 3 are stitched from 2 x 2 confocal images. Scale bars = 100 µm. (B) An example of an anonymized microscope slide with fixed brains used for unbiased scoring under a fluorescent stereomicroscope. Scored brains are marked with a pen to prevent duplicate registration. Scale bar = 500 µm (C) Compared to highly invasive rasV12scrib1 tumors, inhibition of JNK signaling (rasV12scrib1jnkDN) eliminated invasion of clonal cells. Please click here to view a larger version of this figure.
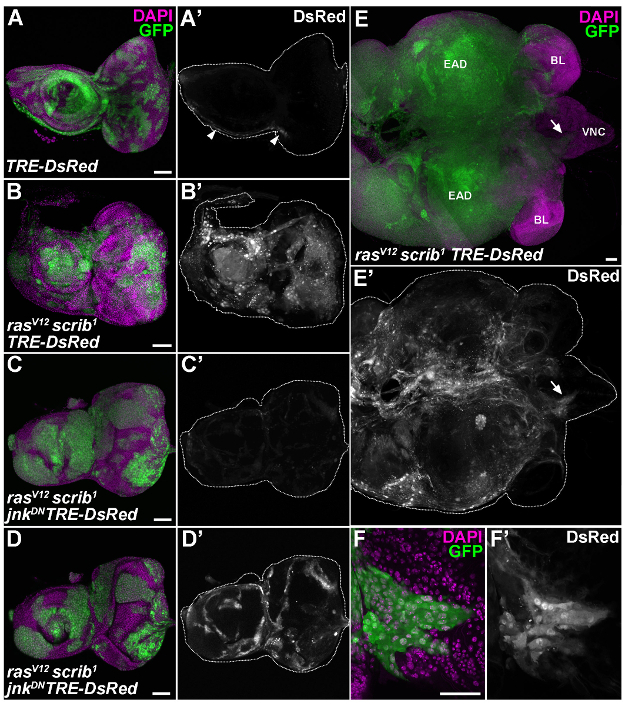
Figure 4: Activation of the transcriptional TRE-DsRed reporter in malignant ras'V12'scrib'1 clonal tumors is JNK-dependent. (A) In control EAD (6 days AEL), the JNK-sensitive TRE-DsRed reporter labeled a narrow stripe of cells running from antennal to eye part (arrowheads). (B) The activity of the TRE-DsRed reporter was markedly enhanced in rasV12scrib1 GFP-positive clones compared to the surrounding non-clonal EAD epithelium. (C) Inhibition of JNK activity resulted in a clear reduction of DsRed signal intensity in rasV12scrib1jnkDN clones. (D) Further enhancement of the DsRed signal revealed non-autonomous activation of JNK signaling in cells surrounding the rasV12scrib1jnkDN clones. (E) On day 8 AEL, rasV12scrib1 cells overgrew the entire EAD and spread over the brain lobes to the VNC (arrow). (F) The clonal cells invading the VNC (close-up of the region marked by the arrow in E) were enriched for the DsRed signal. (A-C) Show projections of multiple confocal sections, (E) is stitched from 3×3 confocal images and (D, F) represent single section. Scale bars = 50 µm. EAD, eye/antennal disc; BL, brain lobe; VNC, ventral nerve cord. Please click here to view a larger version of this figure.
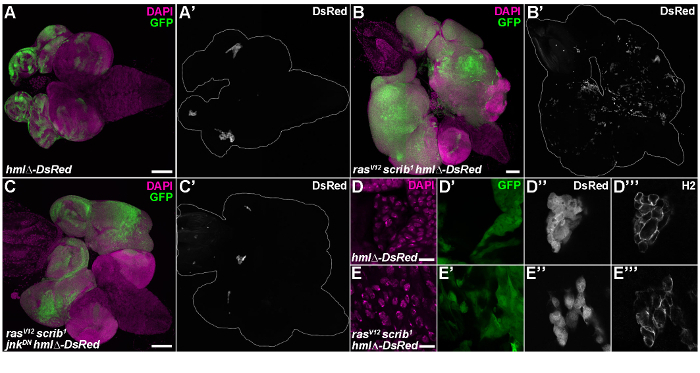
Figure 5: Lineage-specific hmlΔ-DsRed transgenic reporter reveals enrichment of rasV12scrib1 tumor-associated hemocytes. (A) In control EAD, hmlΔ-DsRed reporter-labeled hemocyte clusters are trapped in the indentations of the eye and antennal epithelium. (B) EAD and brain tissue comprised of rasV12scrib1 GFP-marked tumors showed an increased number of hemocytes scattered all over the clonal tissue. (C) The number of associated hemocytes dramatically decreased upon inhibition of JNK signaling in rasV12scrib1jnkDN mosaic tissue. (D-E) HmlΔ-DsRed positive hemocytes were also labeled with an H2-antibody that detects the pan-hemocyte marker Hemese. (A-C) Show projections of multiple confocal sections, (A) is stitched from 2 x 2 and (B-C) are stitched from 3 x 3 confocal images. (D-E) Represent single sections. Scale bars = 100 µm (A-C) and 10 µm (D-E). Please click here to view a larger version of this figure.
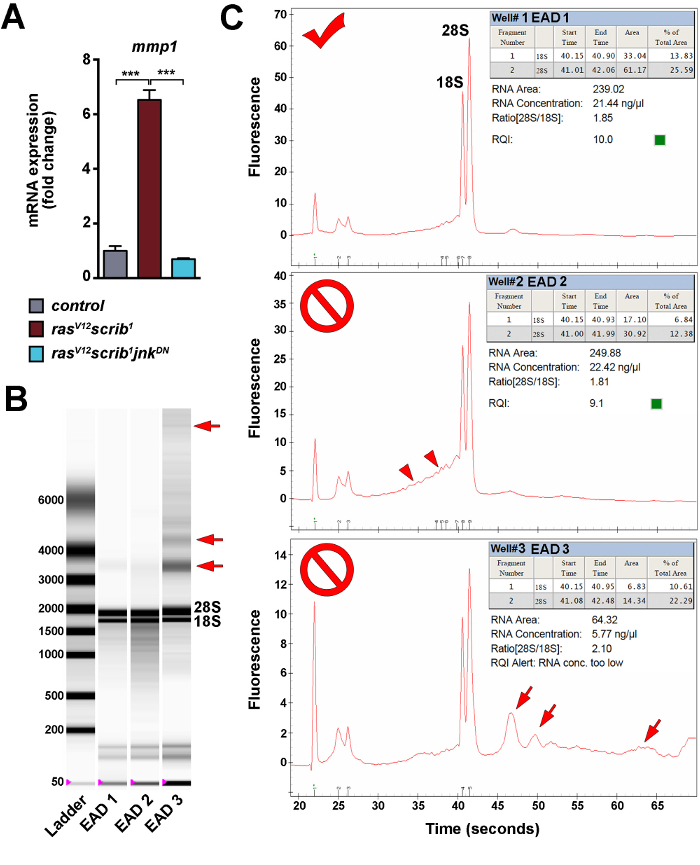
Figure 6: Evaluation of quantity and quality of total RNA isolated from dissected EAD. (A) A representative example of qRT-PCR results using four biological replicates for each genotype. Mmp1 transcript levels were normalized to rp49. Fold changes in gene expression were calculated using the relative standard curve method 32. Data are mean values ± s.e.m.; ***P <0.001 using Student's unpaired two-tailed t-test with unequal variance. (B, C) Assessment of total RNA quality and quantity using an automated electrophoresis system. The virtual gel image (B) shows the two prominent bands of the 18S and 28S rRNAs that correspond to the well-defined peaks on electropherograms (C). DNA contamination and RNA degradation are reflected by a smear (arrowheads) and extranumerary bands and peaks (arrows). Please click here to view a larger version of this figure.
| Fly Strains | Source/Comments |
| eyFLP1; act>y+>Gal4, UAS-GFP; FRT82B, tub-Gal80 | eyFLP-MARCM 82B Green Tester 11 |
| w; UAS-rasV12; FRT82B scrib1/TM6B | 12 |
| w; UAS-rasV12; UAS-jnkDN FRT82B scrib1/TM6B | 12 |
| w; hmlΔ-DsRed; FRT82B | this study, w; hmlΔ-DsRed from Katja Brückner 28 |
| w; UAS-rasV12 hmlΔ-DsRed; FRT82B scrib1/TM6B | this study |
| w; UAS-rasV12 hmlΔ-DsRed; UAS-jnkDN FRT82B scrib1/TM6B | this study |
| w; TRE-DsRed; FRT82B | this study, w; TRE-DsRed from Dirk Bohmann 26 |
| w; UAS-rasV12 TRE-DsRed; FRT82B scrib1/TM6B | this study |
| w; UAS-rasV12 TRE-DsRed; UAS-jnkDN FRT82B scrib1/TM6B | this study |
Table 1: Summary of Drosophila lines used in this study.
