To identify E2 enzymes that cooperate with the ubiquitin ligase CHIP, a set of E2 candidates was tested in individual in vitro ubiquitylation reactions. Cooperating E2-E3 pairs were monitored by the formation of E3-dependent ubiquitylation products, i.e., auto-ubiquitylation of the E3 ligase and the formation of free Ub polymers. The ubiquitylation products were analyzed by western blotting. Data interpretation is based on the size comparison of the resulting protein bands with molecular weight markers. Protein ubiquitylation leads to the formation of specific band patterns characterized by the appearance of double bands or multiple iterative bands with a respective size difference of 8.6 kDa (size of a single Ub molecule).
Here, the ability of nine E2s to promote the formation of ubiquitylation products was tested in the presence of wild-type CHIP (Figure 1A), the inactive U-box mutant CHIP(H260Q) (Figure 1B), or without CHIP (Supplemental Figure S1). E3-independent ubiquitin products were formed in the presence of inactive CHIP and in the absence of CHIP (Figure 1B and Supplemental Figure S1). Inactive CHIP was not auto-ubiquitylated (Figure 1B). In contrast, wild-type CHIP was auto-ubiquitylated when combined with members of the UBE2D family (D1-D3) and members of the UBE2E family (E1, E3), respectively (Figure 1A, lanes 3, 4, and 5). Whereas free poly-Ub chains were produced in cooperation with UBE2D1-3, this was not detected for UBE2E1 or UBE2E3, respectively (Figure 1A, lanes 6 and 7).
The ability of the UBE2D family to promote both the formation of free Ub polymers and the auto-ubiquitylation of CHIP has been attributed to the presence of a non-covalent ubiquitin binding site on the backside of the E227,28. Similarly, the exclusive formation of free Ub chains by UBE2N/V1 (Figure 1A, lane 9) has been attributed to the binding of Ub by a specific UBE2V1 subunit (Uev subunit), directing the formation of K63-linked Ub chains29. No ubiquitylation products were formed in the presence of UBE2C1 and UBE2H (Figure 1A, lanes 2 and 8). In conclusion, CHIP can collaborate with several E2 enzymes in vitro; however, its auto-ubiquitylation is E2 enzyme-specific.
Next, the UBE2D2-CHIP pair was used to investigate the ubiquitylation of the myosin-directed chaperone, UNC-45B, by CHIP ligase activity (Figure 2). Substrate ubiquitylation was analyzed via western blotting. Non-ubiquitylated UNC-45B has a molecular weight of 103 kDa (Figure 2, lane 4). The inactive U-box mutant of CHIP performed neither ubiquitylation of UNC-45B nor auto-ubiquitylation (Figure 2, lane 3). In contrast, ubiquitylation of UNC-45B and auto-ubiquitylation of CHIP was detected upon incubation with wild-type CHIP (Figure 2, lane 6).Thus, CHIP can ubiquitylate UNC-45B in vitro, suggesting that UNC-45B is a conserved substrate of CHIP18.
Ultimately, the catalytic activity of CHIP was analyzed in the absence of any substrate protein. To this end, a lysine discharge assay was performed in which free lysine amino acids can serve as Ub acceptor in the absence of E3 substrates (Figure 3, Supplemental Figure S2C). The discharge assay consists of a charging step in which Ub is loaded onto the E2 by E1, a stop step to prevent further loading of Ub onto E2, and a discharge step where Ub is transferred from the transient Ub~E2 thioester onto free lysine amino acids and/or onto lysine residues of the E3 ligase. Western blot analysis was performed to visualize both the Ub-modified proteins and the E3 ligase CHIP. The uncharged UBE2D2 enzyme has a molecular weight of 17 kDa (Supplemental Figure S2C).
When charged with a single Ub molecule-ensured by the use of lysine-free Ub (Ub K0)-the molecular weight of UBE2D2~Ub shifts upwards to approximately 26 kDa. The time point zero (t0) represents the overall yield of charged E2 (Figure 3). In the presence of inactive CHIP (35 kDa), a faint E3 ligase-independent discharge of UBE2D230,31, but no auto-ubiquitylation of CHIP, was detected. In contrast, in the presence of wild-type CHIP (35 kDa), a faster discharge of UBE2D2 was detected, yielding a complete discharge within 60 min. Simultaneously, auto-ubiquitylation of CHIP was observed, indicating that CHIP promoted the transfer of Ub onto its own lysine residues.
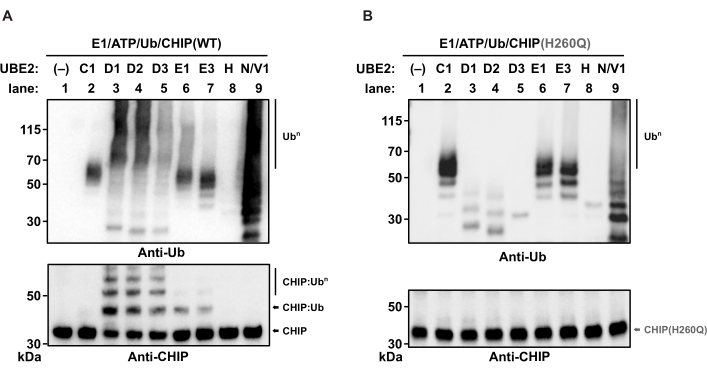
Figure 1: Western blot analysis for E2-E3 enzyme collaboration in vitro. In vitro auto-ubiquitylation reactions with different human E2 enzymes (from left to right: empty, UBE2-D1, -D2, -D3, -E1, -E3, -H, and -N/V1) were performed as indicated using (A) wild-type CHIP or (B) the inactive CHIP U-box mutant, CHIP(H260Q), as E3 ubiquitin ligase. The samples were divided equally, run on separate polyacrylamide gels, and immunoblotted with anti-CHIP and anti-Ub antibodies to visualize reaction products. Abbreviations: Ub = ubiquitin; CHIP = carboxyl terminus of HSC70-interacting protein; ATP = adenosine triphosphate. Please click here to view a larger version of this figure.
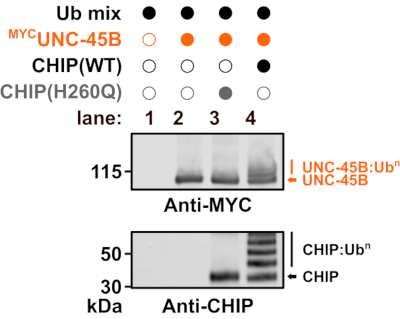
Figure 2: Western blot analysis monitoring substrate ubiquitylation. In vitro substrate ubiquitylation reactions were performed as indicated to detect ubiquitylation of human UNC-45B by CHIP wild-type or the inactive CHIP U-box mutant, CHIP(H260Q). Human UBE2D2 was used as E2 enzyme. Abbreviations: WT = wild-type; Ub = ubiquitin; CHIP = carboxyl terminus of HSC70-interacting protein. Please click here to view a larger version of this figure.
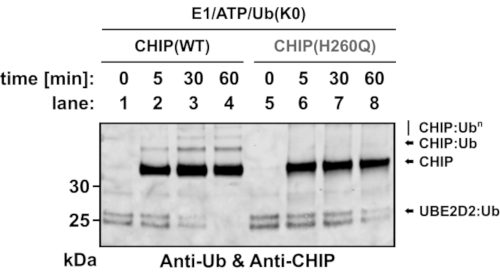
Figure 3: Western blot analysis monitoring the CHIP-dependent Ub transfer from UBE2D2~Ub thioester. Charging of human UBE2D2 by lysine-free Ub (Ub K0), stopping of the charging reaction, and discharging of UBE2D2~Ub was performed as described. For the discharge reactions, wild-type CHIP or the inactive CHIP U-box mutant, CHIP(H260Q), were used as ubiquitin ligases and 10 mM L-lysine was supplied as the potential Ub acceptor. Samples were collected after the indicated time periods, run on a polyacrylamide gel, and immunoblotted with an anti-Ub/anti-CHIP antibody mixture. Abbreviations: Ub = ubiquitin; CHIP = carboxyl terminus of HSC70-interacting protein; ATP = adenosine triphosphate. Please click here to view a larger version of this figure.
Supplemental Figure S1: Representative western blot monitoring E2 enzyme activities in the absence of E3. In vitro ubiquitylation reactions with different human E2 enzymes (from left to right: empty, UBE2-D1, -D2, -D3, -E1, -E3, -H, and -N/V1) were performed as indicated in the absence of an E3 ligase to screen E3-independent reaction products. The samples were run on a polyacrylamide gel and immunoblotted with anti-Ub. Abbreviations: Ub = ubiquitin; ATP = adenosine triphosphate. Please click here to download this File.
Supplemental Figure S2: Analysis of the charging yield and stability of the UBE2D2~Ub K0 thioester. (A) The yield of UBE2D2 charging achieved by E1 was tested under various conditions. The first charging reaction (I) consisted of 5 µM E1, 5 µM E2, and 50 µM Ub K0. The second charging reaction (II) consisted of 2 µM E1, 4 µM E2, and 4 µM Ub K0. The charging reactions were performed as described for 30 min at 37 °C. Samples were collected after 15 and 30 min. The reactions were stopped by addition of 4x LDS sample buffer and incubated at 70 °C for 10 min prior to gel electrophoresis and Coomassie staining. Uncharged UBE2D2 was used as control. Approximately half of the UBE2D2 was converted to UBE2D2~Ub. No additional charging was detected after 15 min. Moreover, neither the increase in E1 nor the increase in free ubiquitin altered the charging yield of UBE2D2~Ub. Thus, charging was subsequently performed for 15 min at 37 °C by using 2 µM E1, 4 µM E2, and 4 µM Ub K0. (B) After charging, the reaction was stopped by addition of 1.8 U/mL apyrase and 30 mM EDTA and incubated at RT. Samples were collected after 2, 5, 8, 10, and 15 min (lanes 2-6), and uncharged UBE2D2 was used as control (lane 1). 4x LDS sample buffer was added, and samples were incubated at 70 °C for 10 min prior to gel electrophoresis and Coomassie staining. Stopping was efficient as measured by the constant band intensity of the UBE2D2~Ub protein, also indicating that the thioester was stable during the indicated time period. (C) Charging of human UBE2D2 by lysine-free Ub (Ub K0), stopping of the charging reaction, and discharging of UBE2D2~Ub was performed as described. For the discharge reactions, wild-type CHIP or the inactive CHIP U-box mutant, CHIP(H260Q) were used as ubiquitin ligases, and 10 mM L-lysine was supplied as the potential Ub acceptor. Samples were collected after the indicated time periods, run on separate polyacrylamide gels, and Coomassie-stained. Please click here download this File.
