ارتفع عدد هياكل الكريستال من البروتينات والمجمعات البروتين بسرعة في السنوات الأخيرة. ما يقدمونه لقطات لا تقدر بثمن للتنظيم الهيكلية لهذه البروتينات وتوفير أساس لتحليل هيكل وظيفة. ومع ذلك، فإن ديناميات البروتينات والتغيرات متعلق بتكوين جزئي، والتي هي ضرورية لمهامهم، ونادرا ما كشفت عنه البلورات بالأشعة السينية. البرد electronmicroscopy، من ناحية أخرى، قادرة على التقاط مجمعات البروتين والبروتين في التشكل مختلفة ولكن بصفة عامة لا يمكن حل التغييرات متعلق بتكوين وصولا الى المستوى الثانوي هيكل 1. ديناميات بتكوين جزئي للبروتينات في الحل في التفاصيل الذرية لا يمكن حلها إلا عن طريق الرنين المغناطيسي، ولكن هذا الأسلوب لا يزال يقتصر على البروتينات ذات أحجام صغيرة نسبيا (عادة ≤ 30 كيلو دالتون) ويحتاج تركيزات عالية من البروتينات (≥ 100 ميكرومتر)، الذي يعيق التجارب مع oligomerization أو تجميع البروتينات عرضة 2. أسلوب واحدقادرة على سد بين عالية الدقة البلورات بالأشعة السينية والبرد electronmicroscopy والذي لا يقتصر حسب حجم البروتين أو تركيز أميد الهيدروجين H-1/2 H-الصرف (HX) في تركيبة مع مطياف الكتلة (MS). في السنوات الأخيرة تطورت هذه الطريقة إلى أداة تحليلية قيمة لتحليل ديناميات البروتين، والبروتين للطي، والاستقرار البروتين والتغيرات متعلق بتكوين 3-5. الأساس الجزيئي لهذه الطريقة هي طبيعة عطوب من الهيدروجين أميد العمود الفقري في البروتينات، والتي سوف تبادل مع ذرات الديوتيريوم عندما يتم وضع البروتين في D 2 O الحل. يتم قياس زيادة لاحقة في البروتين الشامل على مر الزمن مع ارتفاع القرار-MS.
في الببتيدات غير منظم قصيرة HX يعتمد فقط على درجة الحرارة، وتركيز محفز (OH -، H 3 O + أي درجة الحموضة، انظر الشكل 3) وسلاسل الأحماض الأمينية الجانب المخلفات المجاورة بسبب الاستقرائي، والقطآثار alytic والفراغية. هذه التأثيرات الجوهرية على سعر الصرف الكيميائية ك الفصل تم كميا بأناقة من قبل باي وآخرون. 6 والبرنامج متاح (مجاملة Z. تشانغ)، الذي يحسب ك الفصل لكل الأحماض الأمينية داخل ببتيد تعتمد على درجة الحموضة ودرجة الحرارة. في الرقم الهيدروجيني محايدة ودرجات الحرارة المحيطة ك ch غير في الترتيب من 10 1 -10 3 ثانية -1. في البروتينات مطوية HX يمكن أن تكون 2-9 أوامر من حجم أبطأ ويرجع ذلك أساسا إلى الرابطة الهيدروجينية في هيكل الثانوية وبدرجة طفيفة بسبب الوصول المحدود للرطب OH – الأيونات إلى داخل البروتين مطوية بإحكام. لذا HX في البروتينات الأم تورط تتكشف، وتبادل الكيميائية جزئية أو العالمية وطوي ثانية إلى الدولة الأم وفقا للمعادلة (1) وفقا لأسعار الصرف المرصودة ك OBS تعتمد على المرجع سعر الافتتاح ك، سعر الإقفال ك البنود وتبادل الكيميائية الذاتية راالشركة المصرية للاتصالات ك الفصل وفقا لمعادلة (2).
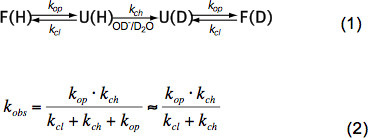
في ظل ظروف الدولة الأم ك المرجع هو أصغر بكثير من الفصل ك ويمكن إهمالها في المقام. هناك نوعان من أنظمة الصرف المدقع دعا EX1 EX2 و. إذا كانت البنود ك هو أصغر بكثير من الفصل ك (EX1) معدل احظ تساوي عمليا إلى سعر الافتتاح وHX يسمح الملاحظة المباشرة للتتكشف من عنصر هيكلي. مثل هذا النظام الصرف، حيث جميع البروتونات أميد الصرف في آن واحد على افتتاح العنصر الهيكلي، ويمكن ملاحظتها بسهولة في MS قبل توزيع ذات النسقين من قمم النظير 7. إذا ك البنود هو أكبر بكثير من الفصل ك (EX2) معدل وحظ يتناسب مع ك الفصل حيث ثابت التناسب يساوي التوازنات-تتكشف للطي المستمر K = ش ك المرجع </sub> / ك البنود. في ظل هذه الظروف، فإن العديد من افتتاح واختتام الأحداث ضرورية قبل كل تبادل البروتونات أميد لدوتيرونس، مما يؤدي إلى زيادة تدريجية في متوسط كتلة بينما يبقى توزيع النظائر تقريبا نفس. النظام EX2 يسمح تحديد الطاقة خالية من تتكشف ΔG u و بالتالي استقرار عنصر هيكلي. تحت شرط الدولة الأم النظام EX2 هو الأكثر شيوعا. زيادة درجة الحموضة وإضافة وكلاء chaotropic يمكن تحويل آلية الصرف لEX1. وبالتالي، HX-MS يمكن استخدامها لاستكشاف الحرارية وكذلك المعلمات الحركية للطي البروتين والتغيرات متعلق بتكوين.
كما ذكر أعلاه HX هو جوهرها ودرجة الحموضة ودرجة الحرارة تعتمد وتبادل عمر النصف للبروتون يتعرض المذيبات تماما من مجموعة أميد العمود الفقري ما بين 5-400 ميللي ثانية في درجة الحموضة الفسيولوجية (الرقم الهيدروجيني 7.6) و 30 درجة مئوية، ولكن 10 دقيقة ل> 15 ساعة بمتوسط> 2 ساعة في درجة الحموضة 2.9 و 0 °C (ما عدا البروتون من أول العمود الفقري أميد السندات من ببتيد، والتي تتبادل مع حياة نصف من كاليفورنيا. 1-2 دقيقة). في ظل هذه الظروف تبادل بطيئة فمن الممكن لهضم العينة باستخدام البروتياز (مثل البيبسين) التي تنشط في ظل هذه الظروف، مع من فقدان جميع المعلومات الواردة في دوتيرونس يدمج. منذ إدخال الهضم المعوية في ظل ظروف تبادل بطيئة، وليس فقط حركية HX العام للبروتينات كامل طول يمكن تحليلها ولكن يمكن أن تكون مترجمة إلى HX مناطق محددة 8،9. يقتصر القرار المكانية حاليا لحجم الشظايا الناتجة الهضمية، والتي هي بشكل عام بين 10-30 المخلفات. ومع ذلك، يمكن تداخل شظايا إنشاؤها نظرا لطبيعة غير محددة من الانقسام قبل البيبسين يؤدي إلى زيادة في القرار المكانية. بالإضافة إلى ذلك، تم العثور على العديد من البروتياز الأخرى أن تكون نشطة في ظل ظروف إخماد، ومع ذلك، أقل كفاءة بكثير من البيبسين 10. مزيد من increaذاتها من القرار المكانية ويمكن الوصول عن طريق تجزئة الببتيدات في الطور الغازي من قبل الأساليب التي حافظت على نمط معالجة بالديوتريوم مثل أسر الإلكترون تفارق (تنمية الطفولة المبكرة)، ونقل الإلكترون تفارق (المفتشون) والأشعة تحت الحمراء multiphoton تفارق (IRMPD) 11-13. هذه التقنيات منع فقدان القرار المكانية بسبب الهجرة داخل الجزيء بروتون ("الهرولة")، والذي لوحظ من قبل الناجم عن الاصطدام تفارق (CID) تقنية تجزئة الأكثر استخداما. ومع ذلك، وهذه الأساليب تتطلب التحسين لكل الببتيد الفردية وبالتالي لا تزال صعبة للغاية.
وقد استخدم HX-MS لتحليل البروتين يجند والبروتين البروتين التفاعلات بما في ذلك التجمع الفيروسية قفيصة 14-17. البروتين تتكشف وطوي ثانية وكذلك تم التحقيق في درجة الحرارة الناجمة عن التغيرات متعلق بتكوين 7،18،19. الفسفرة واحد من الأحماض الأمينية المتصلة بتكوين طفرة يغير 16،20 وnucleotوالتغيرات الناجمة عن بيئة تطوير متكاملة تحليل 21،22. وبالتالي، فإن هذا الأسلوب يبدو مناسبة بشكل مثالي لتحليل التجمع وديناميات الآلات الجزيئية. مرشح واحد جذاب، الذي هو آلية من المصلحة العامة كبيرة، هو مجمع كوصي Hsp90.
